



����1������������������ ������Ĥ�����Ϊ�ǵ��ף���ṹһ���Ϊ�����֣�ϸ��Ĥ�����Ρ���Ĥ���ֺ�ϸ��Ĥ�����Ρ�ϸ��Ĥ�����κ��������ǻ�����ʶ���ز���֮��ϵIJ�λ�����ط��ӺͰ�ϸ������ı��֣���������ԳƵĹ��ܻ��Ź��ɼ�Ϊ���Ӷ��ֿɱ�����幹�͡����غ����������յ����ı䱾���Ĺ�������Ӧ�Է��Ĺ��ͣ����Ϊ���������巢��רһ�Խ���ṩ�����ʻ�����
��������������Ľ������Ϊ������affinity)��һ����˵�����������Ǽ������õĵ�һ�������������뼤�ص�����ѧ��������һ�£������ص���������������϶������ּ��ص����ã��෴ȴ��ϼ������������ϡ�ʵ��֤�����������������������ı仯�������ı䣬�綯�������ڵIJ�ͬ�Σ��ѳ�����ϸ���ϵĵ����ݴ̼��أ�FSH)����������Dz���ͬ�ġ�ijһ������������ʱ�����ڽ����������Ҳ�ɳ������͵�����ҽ.ѧ ȫ��.���ṩwww.med126.com
������������������ı��⣬������Ҳ�ɷ����仯���������ܰ�ϸ��Ĥ���ȵ���������й۲췢�֣��糤��ʹ�ô�������ȵ��أ��������ȵ��������������٣�����Ҳ���ͣ������ȵ��ص������ͺ�����������������ɻָ������������ּ��أ���ټ�״�ټ��ء���ëĤ�����ټ��ء����������ء����ݴ̼��ص�)�������������������ּ���ʹ�������������������ٵ�����Ϊ��˥���ڻ����µ���down regulation0)���µ������Ļ��ƿ����뼤��-���帴������������йء��෴����Щ���أ����ڼ�����Сʱ)Ҳ��ʹ�������������������࣬��Ϊ�������ڻ����ϵ���up regulation)��������ء����ݴ̼��ء�Ѫ�ܽ����صȶ����Գ����ϵ������µ����ϵ�����˵����������ϵ�����˵��������ĺϳ��뽵��ڶ�̬ƽ��֮�У�����������һƽ��Ľ�������Ķ����뼤����������Ӧ���Ե��ڰ�ϸ���Լ������������뷴Ӧǿ�ȡ�
����2��G��������Ϣ�����е����� ���������������ỷ��ø��ϸ��Ĥ������ֿ��ĵ����ʡ����������ϵIJ�����ϸ��Ĥ������棬�����ջ���ø��Ĥ�İ����棬������֮�����һ�����������õĵ��ڵ��ס����������ϵ��ף�guanine nucleotide-binding regulatory protein)�����G���ס�G�����ɦ����ºͦ������ǵ�λ��ɣ����ǵ�λ������������λ�㡣��G�����Ͻ�ϵ�������ΪGTPʱ����������ã�����G�����ϵ�GTPˮ��ΪGDPAʱ��ʧȥ���ԡ���������������ʱ������������G���Ħ��ǵ�λ��ϣ�����ʹ����¡����ǵ�λ���룬���ܶ������ỷ��ø����������á�
����G���ɷ�Ϊ�˷���G���ף�Gs)��������G���� Gi��Gs�������Ǽ��������ỷ��ø���Ӷ�ʹcAMP�������ࣻGi�������������������ỷ��ø�Ļ��ԣ�ʹcAMP���ɼ��١����������ϸ��Ĥ�ļ�������Ҳ�ɷ�Ϊ�˷��ͣ�Rs)�������ͣ�Ri)���֣����Ƿֱ����˷��Լ��أ�Hs)�������Լ���(Hi)������ϣ����ֱ�����Gs��Gi����ͨ����������������ỷ��øʹcAMP���ӻ���ٶ��������á�
����3�������ἡ���Ͷ�������Ϊ�ڶ���ʹ����Ϣ����ϵͳ���ຬ����������cAMPΪ�ڶ���ʹ����ϸ�����ܻ�ģ�����Щ�������ص�������Ϣ������cAMPΪý����д��ݣ����ȵ��ء��߲��ء������ء�ijЩ�����Ե����ĺ��������ӵȡ�ʵ��֤������Щ����������Ĥ�������������ϸ��Ĥ��֬������ת���Ϊ�����ἡ��(inositol-1,4,5,triphosphate,IP3)�Ͷ�������(diacylglycerol,DG)�������°�����Ca2+Ũ�����ߡ����������������IP3��DG�����ǵڶ���ʹ��ѧ˵�������ǵ����ӣ����ҵõ�Խ��Խ�ҵ�ʵ��֤ʵ����һѧ˵��Ϊ���ڼ��ص������£�����ͨ��G���Ľ鵼������ϸ��Ĥ�ڵ���֬øC(phosphinositol-specific phospholipase C.PLC)����ʹ����֬��������PI)�������ữ���ɵ���֬����������PIP2)�ֽ⣬����IP3��DG��DG���ɺ�������Ĥ�У�IP3������������δ�ܵ���������ʱ��ϸ��Ĥ���������������DG��ϸ����IP3�ĺ���Ҳ����ֻ����ϸ��3�ܵ���Ӧ��������ʱ���ż���PIP2�Ľ��⣬��������IP3��DG��IP3�������Ǵ�ʹϸ����Ca2+������ͷ�Ca2+���������ϸ����Ca2+��Ҫ���������������������С�ʵ��֤����IP3����Ca2+���ͷ������������������������壬��Ϊ��������Ĥ����IP3���壬IP3���������������Ϻ���Ca2+ͨ����ʹCa2+���������н��������IP3�շ�Ca2+��Ա�������Ӧ�������ݶ̵��������ͷ�Ca2+���������Ca2+�ͷ��շ����ýϳ���ϸ����Ca2+���������°�����Ca2+Ũ�����ӡ�Ca2+��ϸ���ڵĸƵ����ף�calmodulin,CaM)��Ϻɼ����ø���ٽ����������ữ���Ӷ�����ϸ���Ĺ��ܻ��
����DG��������Ҫ�����������Լ����øC��protein kinase C,PKC)PKC�ļ���������Ca2+�Ĵ��ڡ������PKC��PKAһ����ʹ���ֵ����ʻ�ø�������ữ��Ӧ����������ϸ��������ЧӦ�����⣬DG�Ľ�����ﻨ����ϩ���Ǻϳ�ǰ�����ص�ԭ�ϣ�������ϩ����ǰ�����صĹ��������ֲ��������ỷ��ø�ļ���ٽ�cGMP�����ɡ�CGMP��Ϊ��һ�ֿ��ܵĵڶ���ʹ��ͨ�������øG(PKG)���ı�ϸ���Ĺ��ܣ�ͼ11-3)��

ͼ11-3 ��֬��������Ϣ����ϵͳʾ��ͼ
PIP2����֬�������� DG���������� IP3�������ἡ��PKC������øC CaM:�Ƶ�����
��������)��̴��������û��Ɨ��������ѧ˵
������̴����صķ���С����������Ϊ300����)����֬���ԣ���˿���ϸ��Ĥ����ϸ�����ڽ���ϸ��֮������������Ӱ����������������ã��ʰѴ������û��Ƴ�Ϊ��������ԭ�������Ϊ�������ѧ˵��
������һ���Ǽ�������������ϣ��γɼ���-�������帴����ڰ�ϸ�����д�������̴��������壬�����ǵ����ʣ�����Ӧ���ؽ���ص���רһ��ǿ�����Դ����磬�ӹ���֯�����Ĵƶ�����������17��-�ƶ�����ϣ���������17��-�ƶ�����ϡ���������������Դ�С�뼤�ص�����ǿ����ƽ�еġ����Ұ�������ĺ���Ҳ������ٵĹ���״̬�ı仯�������ı䡣�����ؽ���ϸ��������������Ϻ����嵰�������ͱ仯���Ӷ���������-�������帴�����ý�����ڵ��������ɰ���ת�������ڡ��ڶ�����������������ϣ��γɼ���-�����帴����Ӷ�����DNA��ת¼���̣������µ�mRNA���յ������ʺϳɣ�������Ӧ������ЧӦ��ͼ11-4)��
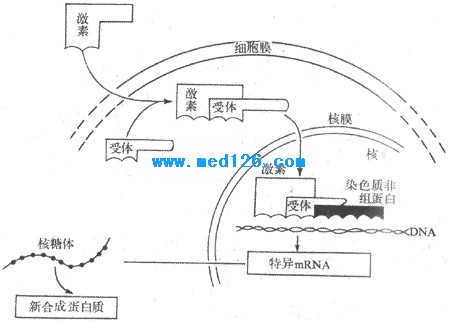
ͼ11-4 ��̴��������û���ʾ��ͼ
�������������ڻ��̼����ķ�չ��Ӧ�ã�������̴����صĺ�������Ľṹ�Ѿ����������������ض�ת¼��������õĵ��ף����������̴����صĿ��ơ���������Ҫ���������ܽṹ���ؽ�Ͻṹ��DNAA�ṹ�ṹ���ת¼��ǿ�ṹ��һ�������������ϣ�����ķ��ӹ������ı䣬��¶�������ڷ����ڲ���DNA��Ͻṹ��ת¼��ǿ�ṹ��ʹ����DNA��ϣ��Ӷ�������ǿת¼��ЧӦ�����⣬���μ�ʵ�����ϱ�������DNA��Ͻṹ�������һ���������е�������Ƭ�ϣ������Ž鵼�������帴������Ⱦɫ�����ض��IJ�λ���ϣ����Ӻ˶�λ�źŵ����á�
������״�ټ��������������أ��������û���ȴ����̴��������ƣ����ɽ���ϸ���ڣ�������������������ϼ�������ڣ���������ϵ��ڻ�����
����Ӧ��ָ�����������ؿ�������ת¼�뷭��ζ�Ӱ�쵰���ʵĺϳɣ�����������̴�����Ҳ����������ϸ��Ĥ����������ѧ˵���Խ��͵�ij��������




