



��������)����ϸ���Ŀ�Ĥ��λ�����γɻ���
������û�������̼�ʱ������ϸ�����ܲ���������λ���������̼������£�����һ�ζ�����λ�������ζ�����λ֮��Ĥ��λ���ȶ�����ġ���������ϸ������������λ3�ڸ���δ�ڴﵽ���ֵ���������λ)֮��4�ڵ�Ĥ��λ�����ȶ�����һˮƽ������������ʼ�Զ��������������е�λ�������˷ܣ�������һ��������λ�����������ܶ���ʼ��������λ�Ͳ��ϵز�����������4�ڵ������Զ��������̣�������ʱ����������ص㣬������ٶ�Զ��0�ڳ�����������ͬ���͵�����ϸ��4�ڳ����ٶȲβһ����ͬ������ϸ��4�ڳ����ٶȱȽϺ㶨������4���Զ����������4�ڻ����������������ڳ���)��������ϸ�������Զ��������˷ܵĻ�����
��������ϸ��Ĥ�����Ŀ�Ĥ�����Ļ������ɿɷ�������ϸ��4���Զ������γɵĻ��ơ������Ʋ⣬����ϸ�����ھ��������ʹĤ������3��)�������λ����4�����ֳ���һ������ǿ�ľ�����������Ӷ�ʹĤ������λ�����ӣ�Ĥ�����������ֽ����Ծ���������IJ��������������ֿ��ܵ�ԭ���������������ǿ���������������˥�ˣ��������С���ͬ���͵�����ϸ����4���Զ��������������ֽ����Ծ�������������𣬵����ɾ�����������������ķ�������ӱ��ʲ�����ȫ��ͬ��
����1���ֿ�Ұϸ���ֿ�Ұϸ����һ�ֿ췴Ӧ����ϸ������Ϊһ�ֿ췴Ӧ��ϸ�������Ķ�����λ����̬�����Ҽ�ϸ�����ƣ����������ӻ���Ҳ������ͬ��
���������ֿ�Ұϸ��4���Զ������γɵĻ��ƣ�80����о����ϱ��������ֿ�Ұϸ�������Ÿ����Ľ��У�����Ĥ����������K+������˥������ͬʱ��Ĥ��λ4�ڿɼ�¼��һ����ʱ�����ƶ�����ǿ�����������If)��ͼ4-7)��Ifͨ���ڶ�����λ3�ڸ�����λ�60mV���ҿ�ʼ������ţ��伤��̶����Ÿ����Ľ��С�Ĥ�ڸ����Ե����Ӷ����ӣ�����100mV���Ҿͳ�ּ����ˣ�����������ֳ�ʱ����������ǿ��Ĥ�ij����̶����Ҳ��ʱ������ӣ�һ���ﵽ�е�λˮƽ�����ֲ�����һ�ζ�����λ�����ͬʱ���������������Ĥ������-50mV������ͨ��ʧ�����ֹ���ɼ���������λ�ĸ�����Ĥ��λ��������������������������ͷ�չ�����أ���������IJ�������ǿ����Ĥ�Ľ����Գ�������Ĥ�ij���һ����������һ�ζ�����λ��һ�����ַ�������ֹ���������������һ�����Ĺ���������ϸ�������ҡ������������ҡ���չ���֡����ҡ����Ƶģ��ɴ˿�������Ϊʲô����ϸ���ܹ��Զ��ء����ϵز����������˷ܡ�

����ͼ4-7 �ֿ�Ұϸ������A����Ĥ��λ B���ɧ�բ�ſ��Ƶ�Ik˥����
�������ɧ�բ�ſ��Ƶ�If���������γ���λ�е���Թ�ϵ
��������4�����������ͨ����Ϊ����������Ҫ���ӳɷ�ΪNa+ ����Ҳ��K+���롣����ʹ����ּ����Ĥ��λΪ-100mV�������Ϊ�������������������һ�ֱ�Ĥ�ij���������ķ�������������Ҫ����Na+)����������־����ΪIf��If��ͨ������Na+ͨ��������ͬ�ڿ�Na+ͨ����������ĵ�ѹˮƽ��ͬ��If�ɱ�藍�Cs)����ϣ������ȴ�����������Ŀǰ������If����ͨ�����о������������ɲ��ܳ�����Խ��͵��ɵ㣬��If�Ľ�һ���о����ܵ��ļ�������ѧ���ǵĸ߶ȹ�ע��
����2����ϸ���Ŀ�Ĥ��λ�����γɻ��� �Ậ�зḻ������ϸ����������λ������������Ե�4���Զ�������������һ������Ӧ����ϸ�������Ĥ��λ�������ͬ�����Ҽ��췴Ӧϸ�����ֿ�Ұ�췴Ӧ����ϸ��������������ϸ���������λ��-70mV)���е�λ��-40mV)�����ڣ���λ����)�ֿ�Ұϸ������0�ڳ�������ʱ��Ĥ�ڵ�λΪ0mV���ң����������Եļ�����ת������������ȣ�70mV)С���ֿ�Ұϸ����Ϊ120mV),��0�ڳ���ʱ�̣�7ms����)ȴ�ֱȺ��ߣ�1-2ms)���öࡣԭ������ϸ��0�ڳ����ٶȣ�Լ10V/s)���������ֿ�Ұϸ����200-1000V/s)����ˣ�������λ��֧Զ���������ô���ͣ���û�����Եĸ���1�ں�ƽ̨�ڣ���4���Զ������ٶȣ�Լ0.1V/s)ȴ���ֿ�Ұϸ����Լ0.02V/s)Ҫ��,��¼��������ϸ��4��Ĥ��λ�仯��б�ʴ����ֿ�Ұϸ��.ͼ4-8��ʾ���Ҽ��췴Ӧϸ������ϸ����Ĥ��λ�仯�IJ��
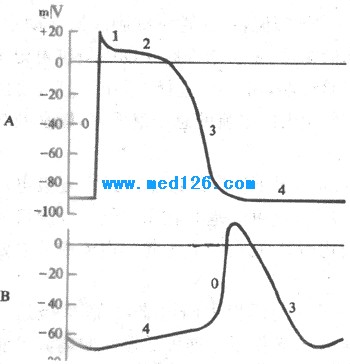
ͼ4-8 ���Ҽ���A)���ᣨB)ϸ����Ĥ��λ�ıȽ�
������ϸ����ֱ����С�����е������о���һ�����ѡ�ֱ��70������ڣ��ſ�ʼ����С�걾�ϲ��õ�ѹǯ���������Ĥ�����������˶����о�����Ŀǰ��δ�ܳ�ֲ������Ŀ�Ĥ��λ��������4�����������ӻ�����ѧ���ǹ۲쵽����ϸ��0�ڳ�������ϸ����Na+Ũ�ȵ�Ӱ�죬�Ժ���ܲ����У��෴������ϸ����Ca2+Ũ�ȵ�����Ӱ�죬���ɱ����Ƹ�ͨ����ҩ������ӣ����첫����D-600��Mn2+��)����ϡ��ݴ˿�����Ϊ��������ϸ��������λ0�ڳ����������������Ca2+���صġ����������������Ϊ�ڶ����������������췴Ӧϸ�������Ҽ����ķ������ֿ�Ұϸ��)0�ڳ����Ŀ�Na+������Ϊ��һ����������������е��о����ϣ��ɽ���ϸ��������λ���γɹ����������£���Ĥ��λ�������λ�Զ��������е�λˮƽʱ������Ĥ�ϸ�ͨ��������Ca2+��������Ica)������0�ڳ��������ͨ����ʧ�Ca2+������Ӧ���٣���һ���棬�ڸ������ڣ���һ��K+ͨ�����������K+��������Ik)��Ca2+���������ٺ�K+�����������ӣ�Ĥ���������ɡ�����ͨ�������ơ���Ca2+����������Ļ���0�ڳ���������ϸ��������λ����Ҫ��������ˣ���Ӧ��Ϊ����Ӧϸ��������Ӧ��λ����������ǰ�����Ҽ��ȿ췴Ӧϸ���Ϳ췴Ӧ��λ��
������ϸ����4���Զ�����Ҳ����ʱ��������ľ�������������𣬵��乹�ɳɷֱȽϸ��ӣ��Ǽ��ֿ�Ĥ�������Ļ�ϡ�Ŀǰ��֪������ϸ��4�ڿ��Լ�¼������Ĥ����������һ����������������������������������ϸ������������õĴ�С�Լ������õ�ʱ��������ͬ��
������1)Ikͨ���ļ��������ǿ����ɵ�K+���������ǵ�����ϸ��������ԭ��Ikͨ����Ĥ������-40mVʱ�㿪ʼ��ʧ�K+������˽������٣�����Ĥ������������Ӷ��γ�4�ڳ�����Ŀǰ��Ϊ������Ikͨ����ʱ����������ʧ������ɵ�K+����������˥��������ϸ��4���Զ�����������Ҫ�����ӻ�����ͼ4-9)��
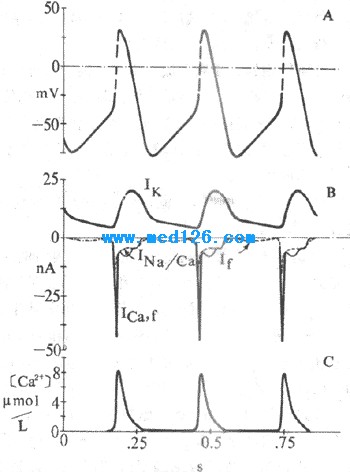
ͼ4-9 �ᶯ����λ����λ�����ӻ���
A����Ĥ��λ B.ԽĤ��λC. ����Ca2+Ũ�ȱ�ʾ������λ��֧��Ica,
f���ɣ���λ ��Ik��If��Ina/Ca����
����(2)If��If��һ�ֽ�������ǿ���������ӣ���ҪΪNa+)�������ֿ�Ұϸ����У�If���ż���Ҫ�����ã���Ik˥�������ú�С�����ǡ�෴����ϸ��4����Ҳ�ɼ�¼��If �����������������ò���Ik˥����ʵ��֤������Cs2+ѡ�������If�����Է���Ƶ�ʽ���ȼ��٣��Լ�����ϸ��4�ھ�����������ܷ�ֵ���ԣ�Ik˥����If�����������õı���Ϊ6��1��
����Ifͨ��������λΪ-100mV���ң��������������ϸ���������λΪ-70mV�������ֵ�λˮƽ�£�Ifͨ���ļ���ʮ�ֻ������������If����4�ڳ����������������ò����ԭ������ϸ������������ʱ��If����ܳ�Ϊ�����е���Ҫ�ɷ֡�
������3)���⣬��ϸ��4���л�����һ�ַ������ԵĻ��������������Ĥ������-60mVʱ������ɼ��������Զ��������̵ĺ�1/3�ڼ�������á����ֻ����������������������Na+-Ca2+�����Ľ����Na+-Ca2+����ʱ���ļ�ϸ���ų�һ��Ca2+������3��Na+����/��ϸ�������֮��Ϊ2��3���γ��������)��
�����ļ�ϸ�������� ���˰��չ��ܺ͵��������Խ��ļ�ϸ����Ϊ����ϸ��������ϸ��֮�⣬�����Ը������������������ر��Ƕ�����λ0�ڳ������ٶȣ����ļ�ϸ����Ϊ�췴Ӧϸ��������Ӧϸ�����䶯����λ��Ӧ��Ϊ�췴Ӧ��λ������Ӧ��λ��Ȼ���ٽ���������ԣ��ɽ��ļ�ϸ����Ϊ�����������ͣ�
�����췴Ӧ������ϸ���������ķ���ϸ�������Ҽ�ϸ����
�����췴Ӧ����ϸ�����ֿ�Ұ����ϸ����
��������Ӧ����ϸ����������ϸ�����Լ��������ͽ�ϣ��������ϸ����
��������Ӧ������ϸ��������ϸ����
�����ļ�ϸ���Ŀ�Ĥ������ �ɶ�������ͨ��ϸ��Ĥ�ϵ������Ի������������ͨ���Լ������ӽ���ת���γɵĿ�Ĥ�����˶������ļ�ϸ����Ĥ��λ�γɵĻ������ļ�ϸ����Ĥ�������������ڶ࣬���ʺܸ��ӣ�������ȡ�������չ��������ʵ���о��ж���Щ����ͨ������������ʶ�Ƚϻ��ң�����ǰ��ì�ܣ��ʽ�Ŀǰ�ȽϿ϶�����Ҫ������������ͨ�������ơ����ڵIJ�λ�����ù��ɽ������£�
����1������������
������1)INa����Ϊ����Na+���������ڿ췴Ӧϸ����������췴Ӧϸ��0�ڳ���������������Ƶ�һ�������)�����ӻ�����
������2)Isi����Ϊ�������������Ҳ�Ƶڶ������������ȥ��Ϊ����һ������ͨ�����ƵĻ���Ca2+�����ʱ�־ΪICa���书���ǹ��ɿ췴Ӧϸ����ƽ̨�ں�����Ӧϸ���ij����ڡ��½����о�����ںܴ�̶��������˶�Ica����ʶ��Ŀǰ��Ϊ���ڶ�������������ǵ�һ��Ca2+�������������Ը����������������ɣ���һ��ֳ�Ica.f����������ֱ��Is1.2��Ica.s�����Т�Ica.f��һ�ֿ���Ca2+ ������ͨ�������ʧ����ٶ�Զ����������Ϊ��Ҫ��öࡣ���ں��ڿ췴Ӧϸ��Na+��������֣���ͬ�γɶ�����λ��֧���϶Σ�����ƽ̨�ڵ����ú�С����������Ӧϸ�����������ӻ������Թ���ϸ�����ԣ�������Ca2+�������ͷ�˧ϸ����Ca2+������������ģ���ȻIca.f�ļ���ʮ��Ѹ�٣���ôCa2+�ͷŵĴ����м��˷ܣ����������������ٶȶ���������Ϊ��Ҫ��öࣻ�ڵ������Ica.s����һ�ֽ�Ica.f����������Ca2+������Ҫ������ά�ֿ췴Ӧϸ��ƽ̨�ڣ���Is1.2�Ƶڶ���������ڶ���֣������ӱ��ʲ��������������Na+-Ca2+�����������������Ҳ��ΪINa.Ca����ƽ̨�������ã�����Ӧ����ϸ�������Զ���������Ҳ���������á�
����(3)If������������ķ���������������������Ҫ��Na+Я��������������ϸ��4�ڡ�If�Ǿ����ֿ�Ұ�췴Ӧ���������Ҫ��������������������Ӧ����ϸ����У������ò���Ik˥����
����2������������
������1)Ikl�������ڿ췴Ӧϸ�����Ǿ����췴Ӧ����ϸ����Ϣ��λ�������������ڸ���2�ں�3�������ã�
������2)Ik���������������Ҫ��K+Я������Ҳ��Na2+�μӣ����ǵ�����K+�������ֳ�Ix���ڿ췴Ӧϸ������3������Ҫ���ã����ֳ�Ϊ�������������Ҳ�������ֿ�Ұϸ���������λ��IxҲ����������Ӧ����ϸ������ʹĤ������4���ڳʽ�����˥�������γ�4�ڳ�������Ҫ���ӻ�����
������3)Ito���ǿ췴Ӧϸ��1�ڸ����������������ӳɷ���ҪΪK+��Ҳ��Na+���롣
����3����ѧ�ſ�����ͨ�� �������֤ʵ�����ļ�ϸ��Ĥ�ϣ����˵�ѹ�ſ�ͨ���⣬�����ڻ�ѧ�ſ�����ͨ�������ǵ����ò�������ά�ֺͲ���������Ĥ��λ����Ϣ��λ�Ͷ�����λ)���������ļ�ϸ����ĵ����Լ��쳣���ĵ����������ر���Ҫ�����塣��Ҫ�Ļ�ѧ�ſ�����ͨ�����������֣�
������1)����������Ƶ�K+ͨ����Ik-ACh)����������ΪACh��������Ikl�����귢����ͨ��G�����������һ��������������ѧ���Ժ�����ѧ�����Ͼ�����ͬ��K+ͨ����
������2)ATP������K+ͨ����Ik-ATP)��ATP�����ò����Ƿֽ�ܼ����ͨ��������ά�ִ�ͨ������������´��ڹر�״̬�����ļ�ϸ����ATP�����ٽ�ˮƽ����ʱ�����ļ�ȱѪʱ)�����������K+ͨ�����š�����K+��©����ȱѪ�ļ�ϸ���ֲ��ض�����������շ�����ʧ����������ҩ�����ϴ�ͨ����
������3)INa�Ck��Ca2+ͨ������ϸ����Ca2+����ʱ�����һ�ַ�������������ͨ��������������Na+��K+���γ�һ����������������Iti)��ʵ���������ijЩ����£��ֿ�Ұϸ���ڶ�����λ������ɲ���һ�ֳ�����λ���ӳٺ������λ)���������е�λʱ�Ϳ����շ���һ���µĶ�����λ���γ���λ������Iti�����ӳٺ������λ�����ӻ��������ػ��ж���ϸ�����K+���Na+���Լ������������Ӱ��ȿ�����ϸ����Ca2+�����ɵ����أ������շ����ǿIti���ӳٺ������λ��




