|
|
||||||||||||
|
|
|||||||||||
|
||||||||||||||||||
| ... | ||
|
||
|
|||||
Пё°ыөДРЛ·ЬРФәНЙъОпөзПЦПу(гРҙМјӨ,ҫІПўөзО»,¶ҜЧчөзО»,KЖҪәвөзО»,·жөзО»,Na+ЖҪәвөзО»,өзС№ЗҜ,ДӨЖ¬ЗҜКөСй,гРөзО»,РЛ·Ьҙ«өј»ъЦЖ ) | |||||
| АҙФҙЈәТҪС§И«ФЪПЯ ёьРВЈә2007-8-25 ТҪС§ВЫМі | |||||
|
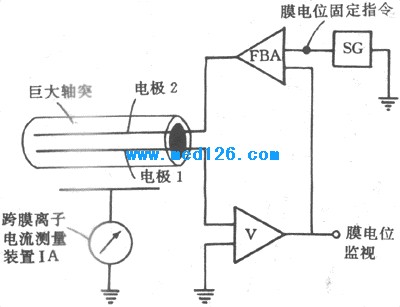
ЎЎЎЎ3.ҫӯөдөДөзС№ЗҜЈЁ»төзС№№М¶ЁЈ©КөСй ҙУЙПКцҝЙЦӘЈ¬HodgkinөИ¶ФУЪ¶ҜЧчөзО»ІъЙъ»ъЦЖөДЛөГчЈ¬№ШјьФЪУЪДӨКЬҙМјӨКұ¶ФNa+ЎўK+өДНЁНёРФ·ўЙъБЛУРСЎФс¶шКұјдТаУРПИәуөДёДұдЈ¬ө«ХвЦ»КЗёщҫЭЛщІвөГөДДӨДЪНвөзО»ёДұд¶ФХХNernst№«КҪҪшРРөДНЖВЫЈ¬КөСйІўГ»УР¶ФДӨөДНЁНёРФҪшРРЦұҪУөДІвБҝәН¶ҜМ¬ГиКцЎЈОӘҙЛЈ¬ЛыГЗУЦУҰУРөұКұЧоПИҪшөДөзЧУС§јјКхЈ¬ЙијЖәНҪшРРБЛЦшГыөДөзС№ЗҜЈЁvoltage clampЈ©КөСйЎЈКөСйөДЙијЖёщҫЭКЗЈәАлЧУЧчҝзДӨТЖ¶ҜКұРОіЙБЛҝзДӨАлЧУөзБчЈЁIЈ©Ј¬¶шНЁНёРФТајҙАлЧУНЁ№эДӨөДДСТЧіМ¶ИЈ¬ҫНКЗДӨөДөзЧиЈЁRЈ©»тЖдө№КэөзөјЈЁGЈ©Ј¬ТтҙЛЛщОҪДӨ¶ФДіЦЦАлЧУНЁНёРФФцҙуКұЈ¬КөјККЗДӨ¶ФёГАлЧУөДөзөјјУҙ󣻶ФУЪҙшөзАлЧУАҙЛөЈ¬ДӨөзөјҫНКЗДӨНЁНёРФөДН¬ТеУпЎЈёщҫЭЕ·Д·¶ЁВЙЈ¬I=VGЈ¬ҝЙЦӘФЪДӨБҪІаөзО»ІоЈЁVЈ©№М¶ЁІ»ұдөДМхјюПВЈ¬ІвіцөДҝзДӨөзБчIөДұд»ҜЈ¬ҫНҝЙЧчОӘДӨөзөјұд»ҜөД¶ИБҝЎЈІв¶ЁДӨФЪКЬҙМјӨКұҝзДӨөзБчөДёДұдФЪјјКхЙПКЗИЭТЧөДЈ¬ө«ФЪХв№эіМЦРТӘұЈіЦДӨөзО»№М¶ЁІ»ұдИҙІ»ИЭТЧЈ»ТтОӘөұҙжФЪҝзДӨАлЧУөзБчКұЈ¬АлЧУөДҪшіцДӨ»бК№І»өјөз¶шУРөзИЭЈЁCЈ©МШРФөДЦ¬ЦКДӨідөз»т·ЕөзЈ¬Тт¶шёщҫЭV=Q/CөД№ШПөЈЁЖдЦРQОӘөзБҝЈ¬ПаөұУЪIәНКұјдtөДіЛ»эЈ©Ј¬ҝзДӨАлЧУөДТЖ¶ҜұШИ»ТӘТэЖрҝзДӨөзО»өДёДұдЈ»КөјКЙПјЗВјөҪөД¶ҜЧчөзО»ҫНКЗХвЦЦёДұдЎЈХэТтОӘИзҙЛЈ¬HodgkinөИЧФРРЙијЖБЛТ»ЦЦУҰУГёә·ҙАЎФӯАнөДөзЧУС§Ч°ЦГЈ¬К№ЛьГЗДЬФЪҝзДӨөзО»О¬іЦәг¶ЁЈЁәг¶ЁөДКэЦөҝЙУЙКөСйХЯНЁ№эКөСйЧ°ЦГФӨПИЙи¶ЁЈ©өДЗйҝцПВЈ¬ІвБҝҝзДӨАлЧУөзБчөДЗҝ¶ИёДұдЈ¬ІўУЙҙЛјЖЛгіцДӨөзөјјҙДӨНЁНёРФөДұд»ҜЗйҝцЎЈөзС№ЗҜКөСйөД»щұҫФӯАнДЈКҪНјИзНј2-12ЛщКҫЎЈНјЦРөзј«1ІеИлҫЮҙуЙсҫӯЦбН»ДЪТ»¶ЁҫаАлЈ¬УГАҙІвБҝәНјаІмХвТ»¶ОЦбН»ДӨДЪөДөзО»Ј¬ҙЛөзј«ПИБ¬өҪТ»ёцөзС№·ЕҙуЖчЈ¬ФЩФЪТ»ёцКҫІЁЖчЙППФКҫЈ»өзј«1ІвөГөзО»Цөҫӯ·ЕҙуәуН¬КұКдёшТ»ёцёә·ҙАЎ·ЕҙуЖчЈЁFBAЈ©Ј¬ХвКЗХыёцТЗЖчЙијЖөД№ШјьІҝ·ЦЈ¬ЛьҝЙ°СІвөГөДДӨДЪөзО»Н¬АҙЧФТ»ёцөзС№ФҙөДЎўУЙКөСйХЯФӨПИЙи¶ЁөДТӘЗуұЈіЦәг¶ЁөДөзО»ЦөҪшРРұИҪПЈ¬Из№ы¶юХЯУРІоЦөЈ¬FBAҫН»бНЁ№эөзј«2ПтЦбН»ДӨДЪКдіцПаУҰЗҝ¶ИәН·ҪПтөДөзБчЈ¬УЙУЪТЗЖчПЯВ·өДҫ«ГЬЙијЖәНҝмЛЩ·ҙУҰЈ¬өзј«2КдіцөзБчөДёДұдХэЧгТФІ№іҘұкұҫУЙУЪҝзДӨАлЧУөзБчК№ДӨід·Еөз¶шТэЖрөДҝзДӨөзО»өДұд¶ҜЈ¬УЪКЗУлөзј«1ПаұЯөДКҫІЁЖчЙППФКҫіцДӨДЪөзО»№М¶ЁФЪЙи¶ЁөДКэЦөЈ¬¶шФЪөзБч·ЕҙуЖчIAЙПІвөГөДҝзДӨАлЧУөзБчөДұд»ҜЈ¬ҫН·ҙУіБЛДӨөзөјөДұд»ҜЎЈ
Нј 2-12 өзС№ЗҜКөСйІјЦГДЈКҪНј ЎЎЎЎөзС№№М¶ЁКөСй»сөГБЛРн¶аУРТвТеөДҪбВЫЎЈКЧПИТ»өгКЗЈ¬Ц»УРЙи¶ЁөДДӨДЪөзО»№М¶ЁФЪИҘј«»ҜЛ®ЖҪКұЈ¬ІЕУРҝЙДЬіцПЦДӨөДNa+өзөјЈЁGNa)әНK+өзөјЈЁGkЈ©өДФцҙуЈ¬ІўЗТЙи¶ЁөзО»УъҪУҪьБгЦөЈ¬өзөјөДФцҙуТІУъГчПФЈ»Па·ҙЈ¬Из№ыЙи¶ЁөДДӨДЪөзО»ЦөКЗі¬ј«»ҜөДЈ¬ФтІ»ҝЙДЬТэЖрҝзДӨАлЧУөзБчәНДӨөзөјөДёДұдЈ¬ХвТ»өгТФәу»№ТӘМёөҪЎЈТФНј2-13өДјЗВјЗъПЯОӘАэЈ¬·ЦОцІ»Н¬АлЧУөДөзөјФЪТ»ҙОРЛ·Ь№эіМЦРөДұд»ҜЗйҝцЎЈНјЦРЧоЙП·ҪЗъПЯұнКҫФЪТ»ҙОөзС№ЗҜКөСйЦРЈ¬°СДӨДЪөзО»УЙҫІПўКұөДЈӯ65mVН»И»№М¶ЁЈЁХвҫНКЗЈЁclampЈ©өДТвЛјЈ©ФЪЈӯ9mVЈ¬Ҫб№ыәЬҝмТэЖрТ»ҙОИзЗъПЯAөДҝзДӨөзБчұд»ҜЗъПЯЈ¬ХвЗъПЯөДҝӘКјІҝ·ЦКЗДЪПтөДЈ¬ТФәуЦрҪҘЧӘұдОӘНвПтөзБчЎЈЦ»јЗВјөҪДЪПт»тНвПтөзБч»№І»ДЬЛөГчөзәЙөДРҜҙшХЯКЗәОЦЦАлЧУЈ¬ёщҫЭ№эИҘөДКөСйХЯУРАнУЙИПОӘЈ¬ПИіцПЦөДДЪПтөзБчҝЙДЬКЗNa+өзБчЈЁINaЈ©Ј¬НвПтөзБчФтҝЙДЬКЗK+өзБчЈЁIkЈ©ЎЈУГёҪјУөДКөСй№ЫІмЦӨГчБЛХвөгЈәјЩ¶Ё°СұкұҫҪюФЎТәЦРөДNaCIУГПаН¬ДҰ¶ыКэөДВИ»ҜөЁјоАҙҙъМжЈ¬ФтФЪН¬СщөДМхјюПВЦ»ДЬјЗВјөҪҪПНніцПЦөДЗъПЯBЈ¬ЛьКЗНвПтөДЈ¬ХвПФИ»КЗТтОӘІ»ДЬіцПЦДЪПтөДINaөДҪб№ыЈ»°СЗъПЯAәНBЦрөгПајхЈ¬ҫНДЬөГөҪЗъПЯCЈ¬ЛьҫНКЗДЪПтөДINaЈ»УЙINaЎўIkБҪМхЗъПЯЈ¬ҫНҝЙЛгіцGNaәНGkөДұд»ҜЗъПЯЈ¬ЖдМШөгКЗЈәЈЁ1Ј©GNaәНGk¶јКЗөзС№ТАҙУРФөДЈ¬Ц»ДЬУЙҝзДӨөзО»өДИҘј«»ҜЛщјӨ»оЈ¬ө«GNaұ»јӨ»оөГФзЈ¬КЗ¶ҜЧчөзО»ЙПЙэЦ§іцПЦөД»щҙЎЈ¬¶шGkјӨ»оіцПЦ»әВэЈ¬КЗ¶ҜЧчөзО»ёҙј«өҪҫІПўөзО»Л®ЖҪөД»щҙЎЈ»ЈЁ2Ј©GNaУРК§»оЈЁinactivationЈ©ЧҙМ¬¶шGkГ»УРҙЛМШРФЈ¬ЖдЦӨГчКЗНј2-13ЦРЗъПЯCЦ»ҙжФЪ1Ў«2msЈ¬ТФәуҝзДӨөзС№ЛдИФ№М¶ЁФЪЈӯ9mVөДЛ®ЖҪЈ¬ө«GNaФзТС»ЦёҙөҪФӯіхЛ®ЖҪЈ¬¶шҙъұнGkөДЗъПЯBЛдИ»іцПЦҪПНнЈ¬ө«ЛьФЪЙи¶ЁөзО»іЦРшЖЪјдТ»ЦұО¬іЦФЪҪПН¬өДЛ®ЖҪЎЈGNaК§»оөДіцПЦәНGkөДјӨ»оКЗФміЙЙсҫӯПЛО¬әН№ЗчАјЎПё°ыұнПЦ¶МҙЩөД·жөзО»өДФӯТтЈ»ФЪДӨёҙј«ТФәуGNaөДК§»оЧҙМ¬ІЕДЬПыК§Ј¬ХвКұGNaІЕДЬТтДӨөДИҘј«»Ҝ¶шФЩіцПЦФцҙуЎЈ
Нј2-13ЎЎөзС№ЗҜКөСйҪб№ыКҫТвНј Ҫ«ҫЮҙуЙсҫӯПЛО¬өДДӨөзО»УЙФӯАҙөД-65mvН»И»ЙПЙэІў№М¶ЁУЪ-9mvөДЛ®ЖҪКұЈ¬ ДӨөДАлЧУөзБчөДұд»ҜЗйҝцЈЁЗъПЯAЎўBЎўCөДТвТејыХэОДЈ© ЎЎЎЎёщҫЭНј2-13ЦРINaәНIkБҪМхөзБчЗъПЯЈ¬јҙҝЙјЖЛгіцН¬ХвБҪХЯПа¶ФУҰөДGNaәНGkЗъПЯЈ¬ФЩёщҫЭХвТ»¶ОДӨЛщҫЯУРөДөзИЭөДКэЦөЈЁУРИЛІвөГГҝcm2өДЗ№ОЪФфЦбН»ДӨөДөзИЭФјОӘ1ҰМFЈ©Ј¬ҫНҝЙЛгіцИз№ыЎ°ФКРнЎұГҝТ»ЛІјдөДАлЧУТЖ¶ҜФЪөзИЭЙПРОіЙөзО»ёДұдКұЈ¬УРҝЙДЬФміЙФхСщөДҝзДӨөзО»өДёДұдЈ¬ХвХэКЗІ»ҪшРРЎ°өзС№№М¶ЁЎұКұөДЗйҝцЈ¬¶шУЙҙЛЧчіцөДөзО»ұд»ҜЗъПЯХэәГН¬ФЪТ»°гКөСйЦРјЗВјөҪөД¶ҜЧчөзО»өДІЁРОМШөгТ»ЦВЈ¬ИзНј2-14ЛщКҫЎЈХвҪшТ»ІҪЛөГчБЛөзС№ЗҜКөСйЦӨГч¶ҜЧчөзО»ІъЙъ»ъЦЖөДХэИ·РФЎЈ ЎЎЎЎ4Ј®ДӨЖ¬ЗҜКөСйәНөҘНЁөААлЧУөзБчөДјЗВјНЁ№эЙПҪЪ№ШУЪөзС№ГЕҝШНЁөАөДМШРФ·ЦОцТСЦӘЈ¬ЛщОҪДӨ¶ФДіЦЦАлЧУНЁНёРФөДёДұдЈ¬КөјКЙПҫц¶ЁУЪДӨҪб№№ЦРУР№ШАлЧУНЁөАө°°ЧЦК·ЦЧУөД№ҰДЬЧҙМ¬Ј»АэИзЈ¬HodgkinөИІвіцөДGNaөДұд»ҜЈ¬КөјККЗДЗТ»¶ОЦбН»ДӨЙПЦЪ¶аөДөзС№ГЕҝШКҪNa+НЁөАТтДӨөДИҘј«»Ҝ¶шҝӘ·ЕөДҪб№ыЎЈФЪHodgkinөИөұКұҪшРРөДДӨөзөјёДұдөДКэС§ДЈДвЦРЈ¬ТСҫӯГчИ·МбКҫЈ¬GNaәНGkөДёДұдІ»КЗҫщФИөШ·ўЙъФЪХыёцДӨЖҪГжЙПЈ¬¶шКЗУлДӨЙПДіР©МШ¶ЁөДЎ°өгЎұУР№ШЈ¬І»ҫГУЦ·ўПЦЈ¬УРР©Т©ОпҝЙТФСЎФсРФөШЧи¶ПДіЦЦАлЧУөДҝзДӨТЖ¶ҜЈ¬ИзәУла¶ҫҝЙТФөҘ¶АЧи¶ПGNa¶шІ»У°ПмGkЈ¬ЛДТТ»щп§ҝЙТФөҘ¶АЧи¶ПGk¶шІ»У°ПмGNaЈ»ТФН¬О»ЛШұкјЗөДәУла¶ҫЦ»ДЬУлДӨЙПДіР©МШКвөДЎ°өгЎұЧчМШТмРФҪбәПЈ¬¶шұкјЗөДЛДТТ»щп§Ц»ДЬУлБнТ»Р©Ў°өгЎұҪбәПЎЈХвР©КөСйТФј°РЛ·Ь№эіМЦРАлЧУТЖ¶ҜКэДҝЦ®¶аУлҝмЈ¬ЦрҪҘК№ИЛГЗНЖ¶ПДӨҪб№№ЦРУРМШКвөДө°°ЧЦКАлЧУНЁөАөДҙжФЪЎЈХвЛөГчЈ¬Ў°НЁөАЎұёЕДоөДМбіцЈ¬Ф¶ФЪНЁөАөДКөЦКұ»ІыГчТФЗ°Ј¬КЗЗ°ХЯҙЩҪшБЛ¶ФәуХЯөДҪшТ»ІҪМҪЛчЎЈ70ДкҙъЦРЖЪУЙNeherәНSakmannөИ·ўХ№іцТ»ЦЦДЬ№»јЗВјДӨҪб№№ЦРөҘТ»өДАлЧУНЁөАө°°ЧЦК·ЦЧУөДҝӘ·ЕәН№ШұХЎўТајҙІвБҝөҘНЁөААлЧУөзБчәНөзөјөДјјКхЈ¬іЖОӘДӨЖ¬ЗҜКөСйЎЈ
Нј 2-14 өзөјұд»ҜУлөзО»ұд»ҜөД№ШПөКҫТвНј ёщҫЭөзС№ЗҜКөСйЦРІвөГөДNa+өзөјЈЁGNaЈ©әНK+өзөјЈЁGkЈ©өДұд»Ҝ№эіМЈ¬ ҝЙТФЛгіцФЪДӨөзО»І»ҪшРРИЛОӘ№М¶ЁКұЈ¬ПаУҰөДNa+ЎўK+АлЧУөзБчФЪДӨөзИЭ ЙПТэЖрөДөзО»ұд»ҜЈЁКөПЯЈ©Ј¬ЖдРОЧҙХэН¬ФЪұкұҫЙПјЗВјөҪөД¶ҜЧчөзО»өДІЁРОТ»ЦВ ЎЎЎЎДӨЖ¬ЗҜКөСйөД»щұҫФӯАнИзНј2-15AЛщКҫЈәУГТ»ёцјв¶Л№вҪаЎўЦұҫ¶Фј0.5Ў«3ҰМmөДІЈБ§Оўөзј«Н¬Йсҫӯ»тјЎПё°ыөДДӨҪУҙҘ¶шІ»ҙМИлЈ¬И»әуФЪОўөзј«БнТ»¶ЛҝӘҝЪК©јУККөұөДёәС№Ј¬Ҫ«Улөзј«јв¶ЛҪУҙҘөДДЗТ»РЎЖ¬ДӨЗб¶ИОьИлөзј«јв¶ЛөДПЛПёҝӘҝЪЈ¬ХвСщФЪХвРЎЖ¬ДӨЦЬұЯУлОўөзј«ҝӘҝЪҙҰөДІЈБ§ұЯСШЦ®јдЈ¬»бРОіЙҪфГЬөД·вҪУЈ¬ФЪАнПлөДЗйҝцПВЖдөзЧиҝЙҙпКэёц»тКэК®З§ХЧЕ·ЈЁЖдОпАн№эіМДҝЗ°ЙРІ»ЗеіюЈ©Ј¬ХвКөјКЙП°СОьёҪФЪОўөзј«јв¶ЛҝӘҝЪҙҰөДДЗТ»РЎЖ¬ДӨН¬ЖдУаІҝ·ЦөДДӨФЪөзС§ЙПНкИ«ёфАлҝӘАҙЈ»Из№ыФЪХвТ»РЎЖ¬ДӨЦРЦ»°ьә¬БЛТ»ёц»тЙЩКэјёёцНЁөАө°°ЧЦК·ЦЧУЈ¬ДЗГҙНЁ№эҙЛОўөзј«ҫНҝЙДЬІвБҝіцөҘТ»НЁөАҝӘ·ЕКұөДАлЧУөзБчәНөзөјЈ¬ІўДЬ¶ФөҘНЁөАөДЖдЛы№ҰДЬМШРФҪшРР·ЦОцЎЈ
Нј2-15 ДӨЖ¬ЗҜКөСйІјЦГКҫТвНј AЈәНјЦРIpОӘјЗВјөҪөДөҘНЁөАөзБчЈ¬VCMDҫц¶ЁЙи¶ЁөДДӨөзО»КэЦө BЈәФЪҙуКуЕЯМҘ№ЗчАјЎПё°ыДӨЖ¬ЙПјЗВјөҪөДУЙACHјӨ»оөДөҘНЁөА ЎЎЎЎҙУNeherөИЧоіхУГДӨЖ¬ЗҜјјКх№ЫІм№ЗчАјЎЦХ°еДӨҙҰөДөҘТ»ACh-ГЕҝШНЁөА»ъДЬМШРФҝӘКјЈ¬ТСҫӯ¶Ф¶аЦЦНЁөАҪшРРБЛ№ЫІмЈ¬·ўПЦЛьГЗТ»°гУРИзПВ№ІН¬МШРФЈәЈЁ1Ј©І»ВЫКЗ»ҜС§ГЕҝШ»төзС№ГЕҝШНЁөАЈ¬ЛьГЗөДҝӘ·ЕәН№ШұХ¶јКЗН»И»өДЈ¬К№Ги»жіцөДөзБчЗъПЯіК·ҪІЁЧҙЈ¬ЛөГчПаУҰөДө°°ЧЦК·ЦЧУҝЙТФҙУТ»ЦЦ№№ПуҝмЛЩөШФҫұдөҪБнТ»ЦЦ№№ПуЈ»ЈЁ2Ј©ГҝЦЦНЁөАҝӘ·ЕКұҫЯУРәг¶ЁөДөзөјЈ¬јҙФЪәг¶ЁөДЗйҝцПВЈ¬Ц»ДЬҝҙөҪЎ°ҝӘЎұ»тЎ°№ШЎұБҪЦЦЧҙМ¬Ј¬әЬЙЩҝҙөҪЎ°°лҝӘЎұ»тЎ°Іҝ·ЦҝӘЎұөДЗйҝцЈ»ЈЁ3Ј©јҙК№КЗН¬Т»НЁөА·ЦЧУЈ¬ГҝҙОҝӘ·ЕөДіЦРшКұјдіӨ¶МТІІ»Т»ЦВЈ¬ЛЖәхЛөГчө°°ЧЦК·ЦЧУҝЙФЪҝӘ·ЕәН№ШұХБҪЦЦ№№ПуЦ®јдЎ°°Ъ¶ҜЎұЈ¬НЈБфФЪДіЦЦЧҙМ¬өДіӨ¶МҫЯУРЛж»ъөДРФЦКЈ»ЈЁ4Ј©ФЪ»ҜС§ГЕҝШНЁөАҪбәПБЛПаУҰөД»ҜС§РЕәЕ·ЦЧУЈ¬»төзС№ГЕҝШНЁөАЛщФЪДӨБҪІаҙҰУЪМШ¶ЁөДөзО»ІоөДЗйҝцПВЈ¬Ў°°Ъ¶ҜЎұөДҙОКэФц¶аЈ¬ҝӘ·ЕөД»ъВКФцҙ󣬶шЎ°К§»оЎұК№ҝӘ·ЕөД»ъВКјхРЎЎЈТҪ.С§И«.ФЪ.ПЯНшХҫwww.lindalemus.com ЎЎЎЎУГөҘНЁөАјЗВјҝЙЛөГчФЪЧФИ»ЗйҝцПВХы¶ОДӨөДАлЧУөзөјәНАлЧУөзБчөДРОіЙ»ъЦЖЈ»ТФЙПКцGNaФцҙуОӘАэЈ¬ЛьПФИ»КЗёГ¶ОДӨЦРЦЪ¶аөДNa+НЁөАФЪИҘј«»ҜөДУ°ПмПВіцПЦҝӘ·ЕөД»ъВКФцјУЛщҫц¶ЁөДЈ¬¶шФЪГҝТ»ЛІјдН¬КұіцПЦөДёчНЁөАөДөзөј»тАлЧУөзБчПа»ҘөюјУЈ¬УЪКЗИзНј2-16BЛщКҫЈ¬ХвЦЦөюјУРОіЙөДNa+өзБчЗъПЯЈ¬ХэәГәННј2-13ЦРөДЗъПЯCПаЛЖЎЈ ЎЎЎЎДӨЖ¬ЗҜКөСйҝЙУГУЪёчЦЦПё°ыЈ¬УЙУЪОўөзј«І»ҙМИлПё°ыЈ¬јҙК№УГУЪПЛРЎөДПё°ыТІІ»ЦВФміЙЛрЙЛЎЈДӨЖ¬ЗҜКөСйТСУРёчЦЦұдКҪЈ¬ИзОьЧЕФЪОўөзј«јв¶ЛөДРЎДӨЖ¬ҝЙТФЛжөзј«¶шН¬ФӯПё°ыНСАлЈ¬°СЛьГЗҪюИлИЛ№ӨҪюФЎТәЦРЈ¬ҫНҝЙТФ№ЫІмДіР©ТтЛШФЪДӨөД°ыҪ¬ІаФхСщУ°ПмНЁөА№ҰДЬЈ»ТІҝЙТФРОіЙДӨөД°ыҪ¬ІаГжПтОўөзј«јв¶ЛҝӘҝЪ¶шДӨұнГжІаГжПтҪюФЎТәөДКөСйДЈКҪЈ¬өИөИЎЈДӨЖ¬ЗҜКөСйТІТСУГУЪПё°ыЙъОпөзТФНвөД№ҰДЬСРҫҝЈ¬ИзПё°ыөД·ЦГЪ№эіМөИЎЈ
Нј2-16 өзС№ГЕҝШNa+НЁөАөДДӨЖ¬ЗҜјЗВјAЈә ЛжЧЕҫІПўөзО»ЈЁEmЈ©УЙЈӯ110mVН»И»№М¶ЁөҪЈӯ50mVЈ¬ ЎЎЎЎИэЎўРЛ·ЬөДТэЖрәНРЛ·ЬөДҙ«өј»ъЦЖ ЎЎЎЎЈЁТ»Ј©гРөзО»әН·жөзО»өДТэЖр ЎЎЎЎДӨДЪёәөзО»ұШРлИҘј«»ҜөҪДіТ»БЩҪзЦөКұЈ¬ІЕДЬФЪХы¶ОДӨТэ·ўТ»ҙО¶ҜЧчөзО»Ј¬ХвёцБЩҪзЦөҙуФјұИХэіЈҫІПўөзО»өДҫш¶ФЦөРЎ10Ў«20mVЈ¬іЖОӘгРөзО»ЎЈАэИзЈ¬ҫЮҙуЙсҫӯЦбН»өДҫІПўөзО»ОӘЈӯ70mVЈ¬ЛьөДгРөзО»ФјОӘЈӯ55mVЎЈХвІ»КЗУЙУЪРЎУЪгРөзО»өДИҘј«»ҜІ»ТэЖрGNaөДФцјУЈ¬КөјКЗйҝцКЗХвКұТІУРТ»¶ЁКэДҝөДNa+НЁөАҝӘ·ЕЈ¬ө«УЙУЪДӨ¶ФK+өДНЁНёРФИФҙуУЪNa+Ј¬Тт¶шЙЩБҝөДNa+ДЪБчј°Жд¶ФДӨДЪөзО»өДУ°ПмЛжјҙұ»K+өДНвБчЛщөЦПыЈ¬Тт¶шИҘј«»ҜІ»ДЬјМРш·ўХ№ПВИҘЈ¬І»ДЬРОіЙ¶ҜЧчөзО»ЎЈЦ»УРөұНвАҙҙМјӨТэЖрөДИҘј«»ҜҙпөҪгРөзО»Л®ЖҪКұЈ¬УЙУЪҪП¶аБҝNa+НЁөАөДҝӘ·ЕФміЙБЛДӨДЪөзО»ҪПҙуөДИҘј«»ҜЈ¬¶шҙЛИҘј«»ҜТСІ»ФЩДЬұ»K+НвБчЛщөЦПыЈ¬Тт¶шДЬҪшТ»ІҪјУҙуДӨЦРNa+НЁөАҝӘ·ЕөД»ъВКЈ¬Ҫб№ыУЦК№ёь¶аNa+ДЪБчФцјУ¶шФміЙДӨДЪҪшТ»ІҪөДИҘј«»ҜЈ¬ИзҙЛ·ҙёҙҙЩҪшЈ¬ҫНРОіЙТ»ЦЦХэ·ҙАЎөД№эіМЈ¬іЖОӘФЩЙъРФСӯ»·Ј¬ЖдҪб№ыК№ДӨДЪИҘј«»ҜСёЛЩ·ўХ№Ј¬РОіЙ¶ҜЧчөзО»¶ёЗНөДЙэЦ§Ј¬ЦұЦБДӨДЪөзО»ЙПЙэөҪҪьУЪNa+ЖҪәвөзО»өДЛ®ЖҪЎЈУЙҙЛҝЙјыЈ¬гРөзО»І»КЗөҘТ»НЁөАөДКфРФЈ¬¶шКЗФЪТ»¶ОДӨЙПДЬК№Na+НЁөАҝӘ·ЕөДКэДҝЧгТФТэЖрЙПГжГиКцөДФЩЙъРФСӯ»·іцПЦөДДӨДЪИҘј«»ҜөДБЩҪзЛ®ЖҪЎЈУЙҙЛТІІ»ДСАнҪвЈ¬Ц»ТӘҙМјӨҙуУЪДЬТэЖрФЩЙъРФСӯ»·өДЛ®ЖҪЈ¬ДӨДЪИҘј«»ҜЛЩ¶ИҫНІ»ФЩҫц¶ЁУЪФӯҙМјӨөДҙуРЎЈ»Хыёц¶ҜЧчөзО»ЙПЙэЦ§өД·щ¶ИТІЦ»ҫц¶ЁУЪФӯАҙҫІПўөзО»өДЦөәНДӨДЪНвөДNa+ЕЁ¶ИІоЈ¬¶шУлТэЖрҙЛҙО¶ҜЧчөзО»өДҙМјӨҙуРЎОЮ№ШЎЈҙЛјҙ¶ҜЧчөзО»ЛщТФДЬұнПЦЎ°И«»тОЮЎұПЦПуөД»ъЦЖЎЈ ЎЎЎЎгРөзО»КЗУГДӨұҫЙнИҘј«»ҜөДБЩҪзЦөАҙГиКц¶ҜЧчөзО»өДІъЙъМхјюЎЈЛщОҪгРЗҝ¶ИЈ¬КЗЧчУГУЪұкұҫКұДЬК№ДӨөДҫІПўөзО»ИҘј«»ҜөҪгРөзО»өДНвјУҙМјӨөДЗҝ¶ИЈ»ХвҫНКЗгРЗҝ¶ИәНгРөзО»ФЪёЕДоЙПөДЗшұрЎЈ ЙПТ»Ті [1] [2] [3] [4] [5] ПВТ»Ті |
|||||
| ОДХВВјИлЈәБиФЖ ФрИОұајӯЈәБиФЖ | |||||
| Ўҫ·ўұнЖАВЫЎҝЎҫјУИлКХІШЎҝЎҫёжЛЯәГУСЎҝЎҫҙтУЎҙЛОДЎҝЎҫ№ШұХҙ°ҝЪЎҝ | |||||
| ЖАВЫҪцҙъұнНшУС№Ыөг,УлұҫХҫОЮ№Ш,ЗлЧсКШПа№Ш·ЁВЙ |
| ЖАВЫјУФШЦР... |
| | ЙиОӘКЧТі | јУИлКХІШ | БӘПөХҫіӨ | RSSөјәҪТіГж | °жИЁЙкГч |НшХҫөШНј | ТҪС§ВЫМі | | |||||
|