|
|
||||||||||||
|
|
|||||||||||
|
||||||||||||||||||
| ... | ||
|
||
|
|||||
Пё°ыөДРЛ·ЬРФәНЙъОпөзПЦПу(гРҙМјӨ,ҫІПўөзО»,¶ҜЧчөзО»,KЖҪәвөзО»,·жөзО»,Na+ЖҪәвөзО»,өзС№ЗҜ,ДӨЖ¬ЗҜКөСй,гРөзО»,РЛ·Ьҙ«өј»ъЦЖ ) | |||||
| АҙФҙЈәТҪС§И«ФЪПЯ ёьРВЈә2007-8-25 ТҪС§ВЫМі | |||||
|
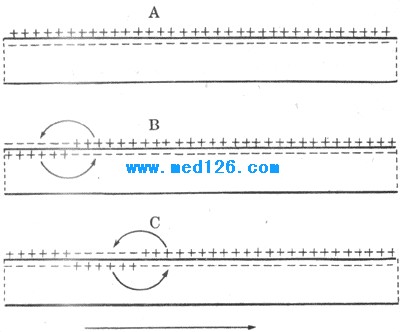
ЎЎЎЎЈЁ¶юЈ©ҫЦІҝРЛ·Ьј°ЖдМШРФ ЎЎЎЎТ»ёцгРПВҙМјӨ»б¶ФҝЙРЛ·ЬПё°ыІъЙъәОЦЦУ°ПмЈҝҝЙНЁ№эНј2-17ЦРөДКөСй»ШҙрЎЈФЪҫЮҙуЙсҫӯЦбН»·ЕЦГТ»¶ФҙМјӨөзј«Ј¬ө«ЖдЦРТ»ёцөзј«ҙ©ИлДӨДЪЈ¬ФЩФЪёҪҪь·ЕЦГТ»ёцЧчДӨДЪөз·ҙУҰјЗВјөДјЗВјөзј«ЎЈјЩ¶ЁПИ°СДӨДЪөДҙМјӨөзј«Б¬өҪөзФҙХэј«Ј¬ДЗГҙөзВ·ҪУНЁКұҪ«»бІъЙъИҘј«»ҜЈ»Из№ыХвёцИҘј«»ҜОҙДЬҙпөҪгРөзО»Ј¬ФтЛөГчЛщУГөзҙМјӨЗҝ¶ИКфУЪгРПВҙМјӨЎЈө«ИзЗ°ЛщКцЈ¬гРПВҙМјӨЛдОҙДЬДӨөзО»ҙпөҪгРөзО»өДИҘј«»ҜЈ¬ТІДЬТэЖрёГ¶ОДӨЦРЛщә¬Na+НЁөАөДЙЩБҝҝӘ·ЕЈ¬Ц»КЗҝӘ·ЕөД»ъВКЙЩЈ¬УЪКЗЙЩБҝДЪБчөДNa+әНөзҙМјӨФміЙөДИҘј«»ҜөюјУЖрАҙЈ¬ФЪКЬҙМјӨөДДӨҫЦІҝіцПЦТ»ёцҪПРЎөДДӨөДИҘј«»Ҝ·ҙУҰЈ¬іЖОӘҫЦІҝ·ҙУҰ»тҫЦІҝРЛ·ЬЈ¬ҫЦІҝРЛ·ЬУЙУЪЗҝ¶ИҪПИхЈ¬ЗТәЬҝмұ»НвБчөДK+ЛщөЦПыЈ¬Тт¶шІ»ДЬТэЖрФЩЙъРФСӯ»·¶ш·ўХ№іЙХжХэөДРЛ·Ь»т¶ҜЧчөзО»ЎЈНј2-17BҫНјЗВјБЛТ»ЧйХвСщөДКөСйЗъПЯЈ¬ЛөГчФЪгРПВҙМјӨөД·¶О§ДЪЈ¬ҙМјӨЗҝ¶ИУъЗҝЈ¬ТэЖрөДДӨөДИҘј«»ҜјҙҫЦІҝРЛ·ЬөД·щ¶ИУъҙуЈЁУЙұнКҫҫІПўөзО»Л®ЖҪөДПЯ¶ОЙП·ҪөДёчМхЗъПЯұнКҫЈ©Ј¬СУРшөДКұјдТІУъіӨЈ»Ц»УРөұҫЦІҝРЛ·ЬөД·щ¶ИҙуөҪЧгТФТэ·ўФЩЙъРФСӯ»·өДЛ®ЖҪКұЈ¬ДӨөДИҘј«»ҜөДЛЩ¶ИІЕН»И»јУҙуЈ¬ХвСщҫЦІҝРЛ·ЬҫН·ўХ№іЙОӘ¶ҜЧчөзО»ЎЈ ЎЎЎЎҫЦІҝРЛ·ЬУРТФПВјёёц»щұҫМШРФЈәЈЁ1Ј©І»КЗЎ°И«»тОЮЎұөДЈ¬¶шКЗЛжЧЕгРПВҙМјӨөДФцҙу¶шФцҙуЈ»ЈЁ2Ј©І»ДЬФЪДӨЙПЧчФ¶ҫаАлөДҙ«ІҘЈ¬ЛдИ»УЙУЪДӨұҫЙнУРөзЧиМШРФЗТДӨДЪНв¶јКЗөзҪвЦКИЬТәЈ¬·ўЙъФЪДӨөДДіТ»өгөДҫЦІҝРЛ·ЬЈ¬ҝЙТФК№БЪҪьөДДӨТІІъЙъАаЛЖөДИҘј«»ҜЈ¬ө«ЛжҫаАлјУҙу¶шСёЛЩјхРЎТФЦБПыК§Ј¬ХвёцҫЦІҝРЛ·ЬЛщІЁј°өД·¶О§ФЪТ»°гЙсҫӯПё°ыДӨЙПІ»і¬№эКэК®ДЛЦБКэ°ЩОўГЧЈ¬ө«УРөДПё°ыұҫЙнТІІ»әЬҙуЈ¬ИзЙсҫӯФӘПё°ыМеЈ¬ҫЦІҝРЛ·ЬөДХвЦЦөзҪфХЕРФА©Іј(eletrotonic propagation)»№КЗУРЦШТӘЙъАнТвТеөДЈ»ЈЁ3Ј©ҫЦІҝРЛ·ЬКЗҝЙТФ»ҘПаөюјУөДЈ¬ТІҫНКЗЛөЈ¬өұТ»ҙҰІъЙъөДҫЦІҝРЛ·ЬУЙУЪөзҪфХЕРФА©ІјЦВК№БЪҪьҙҰөДДӨТІіцПЦіМ¶ИҪПРЎөДИҘј«»ҜЈ¬¶шёГҙҰУЦТтБнТ»ҙМјӨТІІъЙъБЛҫЦІҝРЛ·ЬЈ¬ЛдИ»БҪХЯЈЁөұИ»І»Т»¶ЁПЮУЪБҪХЯЈ©өҘ¶АіцПЦКұ¶јІ»ЧгТФТэ·ўТ»ҙО¶ҜЧчөзО»Ј¬ө«Из№ыУцөҪТ»ЖрКұҝЙТФөюјУЖрАҙЈ¬ТФЦВУРҝЙДЬҙпөҪгРөзО»¶шТэ·ўТ»ҙО¶ҜЧчөзО»ЎЈіЖОӘРЛ·ЬөДҝХјдРФЧЬәНЈ»ҫЦІҝРЛ·ЬөДөюјУТІҝЙТФ·ўЙъФЪБ¬РшКЬКэёцгРПВҙМјӨөДДӨөДДіТ»өгЈ¬ТајҙөұЗ°ГжҙМјӨТэЖрөДҫЦІҝРЛ·ЬЙРОҙПыК§КұЈ¬УләуГжҙМјӨТэЖрөДҫЦІҝРЛ·Ь·ўЙъөюјУЈ¬іЖОӘКұјдРФЧЬәНЎЈЧЬәНПЦПуФЪЙсҫӯФӘПё°ыөД№ҰДЬ»о¶ҜЦРК®·ЦЦШТӘәНіЈјыЎЈБнНвЈ¬УЙНјКҫ2-17BЦР»№ҝЙҝҙөҪЈ¬өұҙМИлДӨДЪөДҙМјӨөзј«әНөзФҙёәј«ПаБ¬КұЈ¬НЁөзКұЦ»ДЬТэЖрДӨөДі¬ј¶»ҜЈЁНјЦРЛ®ЖҪПЯПВ·ҪөДДЗЧйЗъПЯЈ©Ј»ҙМјӨУъЗҝЈ¬і¬ј«»ҜіМ¶ИУъҙуЈ¬ө«І»ТэЖрNa+НЁөАҝӘ·ЕЈ¬ёьІ»ДЬТэ·ў·жөзО»ЎЈКВКөЙПЈ¬ХвКұУЙУЪДӨДЪөзО»әНгРөзО»Ц®јдІоЦөјУҙуЈ¬Тт¶шёГҙҰДӨұдөГёьІ»ИЭТЧРЛ·ЬБЛЎЈМеДЪДіР©ёРКЬЖчПё°ыЎўІҝ·ЦПЩПё°ыәНЖҪ»¬јЎПё°ыЈ¬ТФј°ЙсҫӯПё°ыМеЙПөДН»ҙҘәуДӨәН№ЗчАјЎПё°ыөДЦХ°еДӨЈ¬ЛьГЗФЪКЬҙМјӨКұІ»ІъЙъЎ°И«»тОЮЎұРОКҪөД¶ҜЧчөзО»Ј¬¶шЦ»іцПЦФӯУРҫІПўөзО»өДОўИх¶ш»әВэөДұд¶ҜЈ¬·ЦұріЖОӘёРКЬЖчөзО»ЎўВэөзО»ЎўН»ҙҘәуөзО»әНЦХ°еөзО»ЎЈХвР©өзО»ТІҫЯУРАаЛЖҫЦІҝРЛ·ЬөДМШРФЎЈХвР©РОКҪөДөзұд»ҜЈ¬КөјККЗК№БнТ»Пё°ы»тН¬Т»Пё°ыөДЖдЛыІҝ·ЦөДДӨІъЙъЎ°И«»тОЮЎұКҪ¶ҜЧчөзО»ЙПөД№э¶ЙРФөзұд»ҜЎЈ
Нј2-17 ҫЦІҝРЛ·ЬөДКөСйІјЦГЈЁAЈ©әНКөСйҪб№ыЈЁBЈ©КҫТвНјЛөГчјыХэОД ЎЎЎЎЈЁИэЈ©РЛ·ЬФЪН¬Т»Пё°ыЙПөДҙ«өј»ъЦЖ ЎЎЎЎҝЙРЛ·ЬПё°ыөДМШХчЦ®Т»КЗЛьИОәОТ»ҙҰөДДӨІъЙъөД¶ҜЧчөзО»Ј¬¶јҝЙСШЧЕПё°ыДӨПтЦЬО§ҙ«ІҘЈ¬К№ХыёцПё°ыөДДӨ¶јҫӯАъТ»ҙОАаЛЖУЪұ»ҙМјӨІҝО»өДАлЧУөзөјөДёДұдЈ¬ұнПЦОӘ¶ҜЧчөзО»СШХыёцПё°ыДӨөДҙ«өјЎЈҙ«өјөД»ъЦЖКөјКТС°ьә¬ФЪРЛ·ЬДӨөДЙПКцМШРФЦ®ЦРЎЈЙиПлТ»МхЗ№ОЪФфөДОЮЛиЙсҫӯПЛО¬өДДіТ»РЎ¶ОЈ¬ТтКЬөҪЧг№»ЗҝөДНвјУҙМјӨ¶шіцПЦБЛ¶ҜЧчөзО»ЈЁНј2-18Ј¬BЧу¶ЛЈ©Ј¬јҙёГҙҰіцПЦБЛДӨБҪІаөзО»өДФЭКұРФө№ЧӘЈ¬УЙҫІПўКұөДДЪёәНвХэұдОӘДЪХэНвёәЈ¬ө«әНёГ¶ОЙсҫӯПаБЪҪУөДЙсҫӯ¶ОИФҙҰУЪ°ІҫІКұөДј«»ҜЧҙМ¬Ј»УЙУЪДӨБҪІаөДИЬТә¶јКЗөјөзөДЈ¬УЪКЗФЪТСРЛ·ЬөДЙсҫӯ¶ОәНУлЛьПаБЪөДОҙРЛ·ЬөДЙсҫӯ¶ОЦ®јдЈ¬Ҫ«УЙУЪөзО»ІоөДҙжФЪ¶шУРөзәЙТЖ¶ҜЈ¬іЖОӘҫЦІҝөзБчЎЈЛьөДФЛ¶Ҝ·ҪПтКЗЈәДӨНвУРХэөзәЙУЙОҙРЛ·Ь¶ОТЖПтТСРЛ·Ь¶ОЈ¬ДӨДЪУРХэөзәЙУЙТСРЛ·Ь¶ОТЖПтОҙРЛ·Ь¶ОЎЈХвСщБч¶ҜөДҪб№ыЈ¬КЗФміЙОҙРЛ·Ь¶ОДӨДЪөзО»ЙэёЯ¶шДӨНвөзО»ҪөөНЈ¬ТајҙТэЖрёГҙҰДӨөДИҘј«»ҜЈ»ХвТ»№эіМҝӘКјКұЈ¬ҫНПаөұУЪөзҪфХЕРФА©ІјЎЈёщҫЭЙПКц№ШУЪРЛ·ЬІъЙъөД»ъЦЖөД·ЦОцЈ¬өұИОәОФӯТтК№ДӨөДИҘј«»ҜҙпөҪгРөзО»өДЛ®ЖҪКұЈ¬¶ј»бҙуБҝјӨ»оёГҙҰөДNa+НЁөА¶шөјЦВ¶ҜЧчөзО»өДіцПЦЎЈТтҙЛЈ¬өұҫЦІҝөзБчөДіцПЦК№БЪҪУөДОҙРЛ·ЬөДДӨИҘј«»ҜөҪгРөзО»КұЈ¬ТІ»бК№ёГ¶ОіцПЦЛьЧФјәөД¶ҜЧчөзО»ЎЈЛщОҪ¶ҜЧчөзО»өДҙ«өјЈ¬КөјККЗТСРЛ·ЬөДДӨІҝ·ЦНЁ№эҫЦІҝөзБчЎ°ҙМјӨЎұБЛОҙРЛ·ЬөДДӨІҝ·ЦЈ¬К№Ц®іцПЦ¶ҜЧчөзО»Ј»ХвСщөД№эіМФЪДӨұнГжБ¬РшҪшРРПВИҘЈ¬ҫНұнПЦОӘРЛ·ЬФЪХыёцПё°ыөДҙ«өјЎЈУЙУЪ·жөзО»ІъЙъЖЪјдөзО»ұд»ҜөД·щ¶ИәН¶ё¶ИПаөұҙуЈ¬ТтҙЛФЪөҘТ»Пё°ыҫЦІҝөзБчөДЗҝ¶Иі¬№эБЛТэЖрБЪҪьДӨРЛ·ЬЛщұШРиөДгРЗҝ¶ИКэұ¶ТФЙПЈ¬Тт¶шТФҫЦІҝөзБчОӘ»щҙЎөДҙ«өј№эіМКЗПаөұЎ°°ІИ«ЎұөДЈ¬ТајҙТ»°гІ»ТЧТтДіҙҰ¶ҜЧчөзО»І»ЧгТФК№БЪҪУөДДӨІъЙъРЛ·Ь¶шөјЦВҙ«өјЎ°ЧиЦНЎұЈ¬ХвТ»өгУлТ»°г»ҜС§РФН»ҙҘҙҰөДРЛ·Ьҙ«өЭУРГчПФөДІоұрЎЈ
Нј2-18 ЙсҫӯПЛО¬ҙ«өј»ъЦЖөДДЈКҪНјНдјэН·ұнКҫДӨДЪНв AЈәҫІПўКұ BЈә·ўЙъРЛ·Ьәу CЈәҙ«өј№эіМЦР ЎЎЎЎРЛ·Ьҙ«өј»ъЦЖЛдИ»ТФОЮЛиЙсҫӯПЛО¬ОӘАэЈ¬ө«ФЪЖдЛыҝЙРЛ·ЬПё°ыЈЁИз№ЗчАјЎПё°ыЈ©өДРЛ·Ьҙ«өјЈ¬»щұҫЙПЧсСӯН¬СщөД»ъЦЖЎЈУРЛиЙсҫӯПЛО¬ФЪЦбН»НвГж°ьУРТ»ІгПаөұәсөДЛиЗКЈ¬ЛиЗКЦчТӘіЙ·ЦөДЦ¬ЦККЗІ»өјөз»тІ»ФКРнҙшөзАлЧУНЁ№эөДЈ¬ТтҙЛЦ»УРФЪЛиЗКФЭКұЦР¶ПөДАК·ЙҪбҙҰЈ¬ЦбН»ДӨІЕДЬәНПё°ыНвТәҪУҙҘЈ¬К№ҝзДӨАлЧУТЖ¶ҜөГТФҪшРРЎЈТтҙЛЈ¬өұУРЛиПЛО¬КЬөҪНвјУҙМјӨКұЈ¬¶ҜЧчөзО»Ц»ДЬФЪБЪҪьҙМјӨөгөДАК·ЙҪбҙҰІъЙъЈ¬¶шҫЦІҝөзБчТІЦ»ДЬ·ўЙъФЪПаБЪөДАК·ЙҪбЦ®јдЈ¬ЖдНвөзВ·ТӘНЁ№эЛиЗКНвГжөДЧйЦҜјдТәЈ¬ТтҙЛЈ¬¶ҜЧчөзО»ұнПЦОӘҝз№эГҝТ»¶ОЛиЗК¶шФЪПаБЪАК·ЙҪбҙҰПајМіцПЦЈ¬ХвіЖОӘРЛ·ЬөДМшФҫКҪҙ«өјЎЈ ЎЎЎЎМшФҫКҪҙ«өјКұөДРЛ·Ьҙ«өјЛЩ¶ИЈ¬ПФИ»ұИЙПКцОЮЛиПЛО¬»тТ»°гПё°ыөДҙ«өјЛЩ¶ИҝмөГ¶аЈ»¶шЗТУЙУЪМшФҫКҪҙ«өјКұЈ¬өҘО»іӨ¶ИДЪГҝҙ«өјТ»ҙОРЛ·ЬЛщЙжј°өДҝзДӨАлЧУФЛ¶ҜөДЧЬКэТӘЙЩөГ¶аЈ¬ТтҙЛЛь»№КЗТ»ЦЦЎ°ҪЪДЬЎұөДҙ«өј·ҪКҪЎЈҝҙАҙЈ¬ЙсҫӯЛиЗКөДіцПЦКЗҪш»Ҝ№эіМЦРјИДЬФцјУЙсҫӯПЛО¬ҙ«өјЛЩ¶ИЎўУЦДЬјхЙЩЙъОпДЬБҝПыәДөДТ»ЦЦ·ҪКҪЎЈОЮј№Чө¶ҜОпГ»УРУРЛиЙсҫӯПЛО¬Ј¬¶шОЮЛиПЛО¬ФцјУФцјУҙ«өјЛЩ¶ИөДТ»ёцҝЙДЬНҫҫ¶КЗФцҙуЦбН»өДЦұҫ¶Ј¬ТтОӘХвСщҝЙТФјхЙЩДӨДЪТәМеөДөзЧи¶шФцјУҫЦІҝөзБчөДЗҝ¶ИЈ¬К№¶ҜЧчөзО»өДҙ«өјЛЩ¶ИјУҝмЈ»ХвҙуёЕҫНКЗРиТӘҪшРРҝмЛЩЙсҫӯ·ҙУҰөДЗ№ОЪФфФЪҪш»ҜЦРіцПЦҫЮҙуөДОЮЛиЙсҫӯПЛО¬өДөААнЛщФЪЎЈө«РмҝЖЈЁ1993Ј©өИИЛЦёіцЈ¬ДіР©ОЮј№Чө¶ҜОпөДЙсҫӯПЛО¬ТІҝЙТФТ»ЦЦМШКвөД·ҪКҪҪшРРМшФҫКҪҙ«өјЎЈТҪС§И«ФЪПЯwww.med126.com ЎЎЎЎИз№ыТ»МхЙсҫӯПЛО¬ФЪЛьөДЦРјдІҝКЬөҪҙМјӨЈ¬Ҫ«»бУР¶ҜЧчөзО»УЙЦРјдПтПЛО¬БҪ¶Лҙ«ЛНЈ¬ХвКЗУЙУЪҫЦІҝөзБчҝЙТФіцПЦФЪФӯРЛ·Ь¶ОБҪІаЦ®№КЎЈУЙҙЛҝЙТФАнҪвЈ¬РЛ·ЬФЪН¬Т»Пё°ыЙПөДҙ«өјЈ¬ІўІ»ПЮУЪіҜПтДіТ»·ҪПтЈ»МеДЪЙсҫӯПЛО¬ЛщТФУРҙ«ИләНҙ«іцЦ®·ЦЈ¬Ц»КЗУЙУЪФЪХыМеөДЧФИ»МхјюПВЈ¬ҙ«ИлПЛО¬Ц»ДЬФЪЛьГЗәНёРКЬЖчПаБ¬ҪУөДНвЦЬ¶Лұ»ҙМјӨЈ¬¶шҙ«іцПЛО¬Ц»ДЬФЪЛьГЗөДПё°ыМеІъЙъіе¶Ҝ¶шҙ«ПтНвЦЬЈ¬Іў·ЗКЗУЙУЪХвР©ПЛО¬ұҫЙнЦ»ДЬөҘ·ҪПтҙ«өјРЛ·ЬөДФө№КЎЈТФ¶ҜЧчөзО»ОӘРЛ·ЬіцПЦөДЦёұкЈ¬ҝЙТФІв¶ЁРЛ·ЬФЪёчЦЦПё°ыөДҙ«өјЛЩ¶ИЎЈАэИзЈ¬ИЛМеТ»Р©ҪПҙЦөДУРЛиЙсҫӯПЛО¬өДҙ«өјЛЩ¶ИЈ¬ЧоҝмҝЙҙпГҝГл100mТФЙПЈ¬¶шТ»Р©Пё°ыөДОЮЛиПЛО¬ГҝГлҙ«өјҫаАл»№І»өҪ1mЈ»№№іЙРДФаДЪІҝҙ«өјПөНіөДЖЦҝПТ°Пё°ыЈ¬ГҝГлҙ«өјЛЩ¶ИФј4Ў«5mЈ¬КЗРДјЎПё°ыЦРҙ«өјЛЩ¶ИЧоҝмөДЎЈ |
|||||
| ОДХВВјИлЈәБиФЖ ФрИОұајӯЈәБиФЖ | |||||
| Ўҫ·ўұнЖАВЫЎҝЎҫјУИлКХІШЎҝЎҫёжЛЯәГУСЎҝЎҫҙтУЎҙЛОДЎҝЎҫ№ШұХҙ°ҝЪЎҝ | |||||
| ЖАВЫҪцҙъұнНшУС№Ыөг,УлұҫХҫОЮ№Ш,ЗлЧсКШПа№Ш·ЁВЙ |
| ЖАВЫјУФШЦР... |
| | ЙиОӘКЧТі | јУИлКХІШ | БӘПөХҫіӨ | RSSөјәҪТіГж | °жИЁЙкГч |НшХҫөШНј | ТҪС§ВЫМі | | |||||
|