|
|
||||||||||||
|
|
|||||||||||
|
||||||||||||||||||
| ... | ||
|
||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
жзСіЕФВЁвђбЇКЭЗЂВЁбЇ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| РДдДЃКвНбЇШЋдкЯп ИќаТЃК2007-8-26 вНбЇТлЬГ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
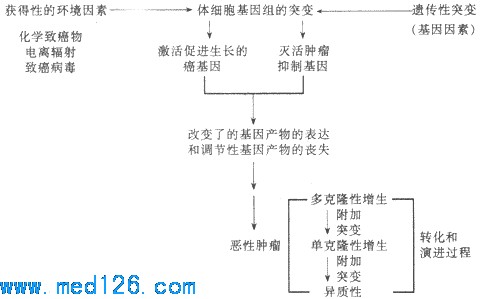
жзСіВЁвђбЇбаОПв§Ц№жзСіЕФЪМЖЏвђЫиЃЌжзСіЗЂВЁбЇдђбаОПжзСіЕФЗЂВЁЛњжЦгыжзСіЗЂЩњЕФЬѕМўЁЃвЊжЮгњжзСіКЭдЄЗРжзСіЕФЗЂЩњЃЌЙиМќЮЪЬтЪЧВщУїжзСіЕФВЁвђМАЦфЗЂВЁЛњжЦЁЃ ЁЁЁЁЙигкжзСіЕФВЁвђбЇКЭЗЂВЁбЇЃЌЖрФъРДНјааСЫЙуЗКЕФбаОПЃЌЫфШЛжСНёЩаЮДЭъШЋВћУїЃЌЕЋНќФъРДЗжзгЩњЮябЇЕФбИЫйЗЂеЙЃЌЬиБ№ЪЧЖдАЉЛљвђКЭжзСівжжЦЛљвђЕФбаОПЃЌвбОГѕВННвЪОСЫФГаЉжзСіЕФВЁвђгыЗЂВЁЛњжЦЃЌР§ШчВЎЛљЬиСмАЭСіКЭШЫРрTЯИАћАзбЊВЁ/СмАЭСіЁЃФПЧАЕФбаОПБэУїЃЌжзСіДгБОжЪЩЯЫЕЪЧЛљвђВЁЁЃв§Ц№вХДЋЮяжЪDNAЫ№КІЃЈЭЛБфЃЉЕФИїжжЛЗОГЕФгывХДЋЕФжТАЉвђзгПЩФмвдаЭЌЕФЛђепађЙсЕФЗНЪНЃЌМЄЛюАЉЛљвђЛђЃЈКЭЃЉУ№ЛюжзСіЕФвжжЦЛљвђЃЌЪЙЯИАћЗЂЩњзЊЛЏЃЈtransformationЃЉЁЃБЛзЊЛЏЕФЯИАћПЩЯШГЪЖрПЫТЁаддіЩњЃЌОЙ§вЛИіТўГЄЕФЖрНзЖЮЕФбнНјЙ§ГЬЃЈprogressionЃЉЃЌЦфжавЛИіПЫТЁПЩЯрЖдЮоЯожЦЕиРЉдіЃЌЭЈЙ§ИНМгЭЛБфЃЌбЁдёадЕиаЮГЩОпгаВЛЭЌЬиЕуЕФбЧПЫТЁЃЈвьжЪадЃЉЃЌДгЖјЛёЕУНўШѓКЭзЊвЦЕФФмСІЃЈЖёадзЊЛЏЃЉЃЌаЮГЩЖёаджзСіЁЃЭМ7Ѓ19ЪОжзСіЕФВЁвђКЭЗЂВЁЛњжЦФЃЪНЁЃ
ЭМ7Ѓ19жзСіЕФВЁвђКЭЗЂВЁЛњжЦФЃЪНЭМЃЈВЩздKumarЃЌЩдИФЃЉ ЁЁЁЁвЛЁЂжзСіЗЂЩњЕФЗжзгЩњЮябЇЛљДЁ ЁЁЁЁ1.АЉЛљвђ ЁЁЁЁЃЈ1ЃЉдАЉЛљвђЁЂАЉЛљвђМАЦфВњЮяЃКЯжДњЗжзгЩњЮябЇЕФжиДѓГЩОЭжЎвЛЪЧЗЂЯжСЫдАЉЛљвђЃЈproto-oncogeneЃЉКЭдАЉЛљвђОпгазЊЛЏГЩжТАЉЕФАЉЛљвђЃЈoncogeneЃЉЕФФмСІЁЃBishopКЭVarmusвђЮЊдкетЗНУцЕФЙБЯзЖјЛёЕУ1989ФъЕФХЕБДЖћНБЁЃ ЁЁЁЁАЉЛљвђЪЧЪзЯШдкФцзЊТМВЁЖОЃЈRNAВЁЖОЃЉжаЗЂЯжЕФЁЃКЌгаВЁЖОАЉЛљвђЕФФцзЊТМВЁЖОФмдкЖЏЮябИЫйгеЗЂжзСіВЂФмдкЬхЭтзЊЛЏЯИАћЁЃКѓРДдке§ГЃЯИАћЕФDNAжавВЗЂЯжСЫгыВЁЖОАЉЛљвђМИКѕЭъШЋЯрЭЌЕФDNAађСаЁЃБЛГЦЮЊЯИАћАЉЛљвђЁЃШчras,mycЕШЁЃгЩгкЯИАћАЉЛљвђдке§ГЃЯИАћжаФЫвдЗЧМЄЛюЕФаЮЪНДцдкЃЌЙЪгжГЦЮЊдАЉЛљвђЁЃдАЉЛљвђПЩвдгЩгкЖржжвђЫиЕФзїгУЪЙЦфНсЙЙЗЂЩњИФБфЃЌЖјБЛМЄЛюГЩЮЊАЉЛљвђЁЃ ЁЁЁЁдАЉЛљвђБрТыЕФЕААзжЪДѓЖрЪЧЖде§ГЃЯИАћЩњГЄЪЎЗжживЊЕФЯИАћЩњГЄвђзгКЭЩњГЄвђзгЪмЬхЃЌШчбЊаЁАхбмЩњЩњГЄвђзгЃЈPDGFЃЉЁЂЯЫЮЌФИЯИАћЩњГЄвђзгЃЈFGFЃЉЁЂБэЦЄЯИАћЩњГЄвђзгЪмЬхЃЈEGFЃRЃЉЁЂживЊЕФаХКХзЊЕМЕААзжЪЃЈШчРвАБЫсМЄУИЁЂЫПАБУИЃЫеАБЫсМЄУИЕШЃЉвдМАКЫЕїНкЕААзЃЈШчзЊТММЄЛюЕААзЃЉЕШЁЃБэ7Ѓ4ЪОГЃМћЕФАЉЛљвђМАЦфВњЮяЁЃ Бэ7Ѓ4 МИжжГЃМћЕФАЉЛљвђМАЦфМЄЛюЗНЪНКЭЯрЙиЕФШЫРржзСі
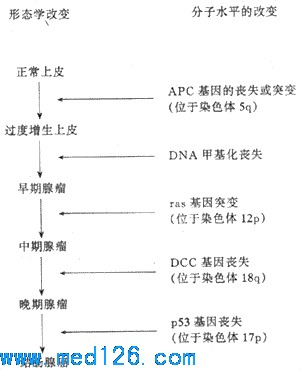
ЃЈВЩздBasic PathologyЃЛ1992ЃЉ ЁЁЁЁЃЈ2ЃЉдАЉЛљвђЕФМЄЛюЃКдАЉЛљвђдкИїжжЛЗОГЕФЛђвХДЋЕФвђЫизїгУЯТЃЌПЩЗЂЩњНсЙЙИФБфЃЈЭЛБфЃЉЖјБфЮЊАЉЛљвђЃЛвВПЩвдЪЧдАЉЛљвђБОЩэНсЙЙУЛгаИФБфЃЌЖјЪЧгЩгкЕїНкдАЉЛљвђБэДяЕФЛљвђЗЂЩњИФБфЪЙдАЉЛљвђЙ§ЖШБэДяЁЃвдЩЯЛљвђЫЎЦНЕФИФБфПЩМЬЖјЕМжТЯИАћЩњГЄДЬМЄаХКХЕФЙ§ЖШЛђГжајГіЯжЃЌЪЙЯИАћЗЂЩњзЊЛЏЁЃв§Ц№дАЉЛљвђЭЛБфЕФDNAНсЙЙИФБфАќРЈЕуЭЛБфЃЈШч90ЃЅЕФвШЯйАЉгаras ЛљвђЕФЕуЭЛБфЃЉЁЂШОЩЋЬхвзЮЛЃЈШчВЎЛљЬиСмАЭСіЕФt(8ЃК14)ЃЌТ§адСЃЯИАћАзбЊВЁЕФPhlШОЩЋЬхЃЉЁЂВхШыгеБфЁЂЛљвђШБЪЇКЭЛљвђРЉдіЃЈШчЩёОФИЯИАћСіЕФNЃmycдАЉЛљвђПЩИДжЦГЩЖрДяМИАйИіПНБДЃЌдкЯИАћвХДЋбЇЩЯБэЯжЮЊШОЩЋЬхГіЯжЫЋЮЂаЁЬхКЭОљШОЧјЃЉЁЃАЉЛљвђБрТыЕФЕААзжЪЃЈАЉЕААзЃЉгыдАЉЛљвђЕФе§ГЃВњЮяЯрЫЦЃЌЕЋгажЪЛђСПЕФВЛЭЌЁЃЭЈЙ§ЩњГЄвђзгЛђЩњГЄвђзгЪмЬхдіМгЁЂВњЩњЭЛБфЕФаХКХзЊЕМЕААзгыDNAНсКЯЕФзЊТМвђзгЕШЛњжЦЃЌАЉЕААзЕїНкЦфАаЯИАћЕФДњаЛЁЂДйЪЙИУЯИАћж№ВНзЊЛЏЃЌГЩЮЊжзСіЯИАћЁЃ ЁЁЁЁ2ЃЎжзСівжжЦЛљвђЁЁгыдАЉЛљвђБрТыЕФЕААзжЪДйНјЯИАћЩњГЄЯрЗДЃЌдке§ГЃЧщПіЯТДцдкгкЯИАћФкЕФСэвЛРрЛљвђЁЊЁЊжзСівжжЦЛљвђЕФВњЮяФмвжжЦЯИАћЕФЩњГЄЁЃШєЦфЙІФмЩЅЪЇдђПЩФмДйНјЯИАћЕФжзСіадзЊЛЏЁЃгЩДЫПДРДЃЌжзСіЕФЗЂЩњПЩФмЪЧАЉЛљвђЕФМЄЛюгыжзСівжжЦЛљвђЕФЪЇЛюЙВЭЌзїгУЕФНсЙћЁЃФПЧАСЫНтзюЖрЕФСНжжжзСівжжЦЛљвђЪЧRbЛљвђКЭP53ЛљвђЁЃЫќУЧЕФВњЮяЖМЪЧвдзЊТМЕїНквђзгЕФЗНЪНПижЦЯИАћЩњГЄЕФКЫЕААзЁЃЦфЫќжзСівжжЦЛљвђЛЙгаЩёОЯЫЮЌСіВЁЃ1ЛљвђЁЂНсГІЯйСіадЯЂШтЛљвђЁЂНсГІАЉЖЊЪЇЛљвђКЭWilmsСіЃ1ЕШЁЃ ЁЁЁЁЃЈ1ЃЉRbЛљвђЃКRbЛљвђЫцзХЖдвЛжжЩйМћЕФЖљЭЏжзСіЁЊЁЊЪгЭјФЄФИЯИАћСіЕФбаОПЖјзюдчЗЂЯжЕФвЛжжжзСівжжЦЛљвђЁЃRbЛљвђЕФДПКЯзгадЕФЖЊЪЇМћгкЫљгаЕФЪгЭјФЄФИЯИАћСіМАВПЗжЙЧШтСіЁЂШщЯйАЉКЭаЁЯИАћЗЮАЉЕШЁЃRbЛљвђЖЈЮЛгкШОЩЋЬх13q14ЃЌБрТывЛжжКЫНсКЯЕААзжЪЃЈP105ЃRbЃЉЁЃЫќдкЯИАћКЫжавдЛюЛЏЕФЭбСзЫсЛЏКЭЪЇЛюЕФСзЫсЛЏЕФаЮЪНДцдкЁЃЛюЛЏЕФRbЕААзЖдгкЯИАћДгG0/G1ЦкНјШыSЦкгавжжЦзїгУЁЃЕБЯИАћЪмЕНДЬМЄПЊЪМЗжСбЪБЃЌRb ЕААзБЛСзЫсЛЏЪЇЛюЃЌЪЙЯИАћНјШыSЦкЁЃЕБЯИАћЗжСбГЩСНИізгЯИАћЪБЃЌЪЇЛюЕФЃЈСзЫсЛЏЕФЃЉRbЕААзЭЈЙ§ЭбСзЫсЛЏдйЩњЪЙзгЯИАћДІгкG1ЦкЛђG0ЕФОВжЙзДЬЌЁЃШчЙћгЩгкЕуЭЛБфЛђ13q14ЕФЖЊЪЇЖјЪЙRbЛљвђЪЇЛюЃЌдђRbЕААзЕФБэДяОЭЛсГіЯжвьГЃЃЌЯИАћОЭПЩФмГжајЕиДІгкдіжГЦкЃЌВЂПЩФмгЩДЫЖёБфЁЃ ЁЁЁЁЃЈ2ЃЉp53ЛљвђЃКp53ЛљвђЖЈЮЛгк17КХШОЩЋЬхЁЃе§ГЃЕФp53ЕААзЃЈвАЩњаЭЃЉДцдкгкКЫФкЃЌдкЭбСзЫсЛЏЪБЛюЛЏЃЌгазшАЯИАћНјШыЯИАћжмЦкЕФзїгУЁЃдкВПЗжНсГІАЉЁЂЗЮАЉЁЂШщЯйАЉКЭвШЯйАЉЕШОљЗЂЯжгаp53ЛљвђЕФЕуЭЛБфЛђЖЊЪЇЃЌДгЖјв§Ц№вьГЃЕФp53ЕААзБэДяЃЌЖјЩЅЪЇЦфЩњГЄвжжЦЙІФмЃЌДгЖјЕМжТЯИАћдіЩњКЭЖёБфЁЃНќРДЛЙЗЂЯжФГаЉDNAВЁЖОЃЌР§ШчHPVКЭSVЃ40ЃЌЦфжТАЉзїгУЪЧЭЈЙ§ЫќУЧЕФАЉЕААзгыЛюЛЏЕФRbЕААзЛђp53ЕААзНсКЯВЂжаКЭЦфЩњГЄвжжЦЙІФмЖјЪЕЯжЕФЁЃ ЁЁЁЁ3ЃЎЖрВНАЉБфЕФЗжзгЛљДЁЁЁЖёаджзСіЕФЗЂЩњЪЧвЛИіГЄЦкЕФЁЂЖрвђЫидьГЩЕФЗжНзЖЮЕФЙ§ГЬЃЌетвбгЩСїааВЁбЇЁЂвХДЋбЇКЭЛЏбЇжТАЉЕФЖЏЮяФЃЪНЫљжЄУїЁЃНќФъРДЕФЗжзгвХДЋбЇбаОПДгАЉЛљвђКЭжзСівжжЦЛљвђЕФНЧЖШЮЊДЫЬсЙЉСЫИќМггаСІЕФжЄУїЁЃЕЅИіЛљвђЕФИФБфВЛФмдьГЩЯИАћЕФЭъШЋЖёадзЊЛЏЃЌЖјЪЧашвЊЖрЛљвђЕФИФБфЃЌАќРЈМИИіАЉЛљвђЕФМЄЛюКЭСНИіЛђИќЖржзСівжжЦЛљвђЕФЩЅЪЇЁЃвдНсГІАЉЕФЗЂЩњЮЊР§ЃЌдкДгНсГІЩЯЦЄЙ§ЖШдіЩњЕННсГІАЉЕФбнНјЙ§ГЬжаЃЌЙиМќадЕФВНжшЪЧАЉЛљвђвдМАжзСівжжЦЛљвђЕФЩЅЪЇЛђЭЛБфЁЃетаЉНзЖЮЬнадЛ§РлЦ№РДЕФВЛЭЌЛљвђЗжзгЫЎЦНЕФИФБфЃЌПЩвддкаЮЬЌбЇЕФИФБфЩЯЗДгГГіРДЃЈЭМ7Ѓ20ЃЉЁЃ
ЭМ7Ѓ20ЁЁНсжБГІАЉЭЈЙ§ЩЯЦЄдіЩњЃЯйСіЃАЉЕФНзЬнадЕФбнНјЕФЗжзгЩњЮябЇКЭаЮЬЌбЇИФБфЕФЙиЯЕ ЃЈВЩздKumarЃЉ ЁЁЁЁ |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ЮФеТТМШыЃКСшдЦ д№ШЮБрМЃКСшдЦ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ЁОЗЂБэЦРТлЁПЁОМгШыЪеВиЁПЁОИцЫпКУгбЁПЁОДђгЁДЫЮФЁПЁОЙиБеДАПкЁП | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ЦРТлНіДњБэЭјгбЙлЕу,гыБОеОЮоЙи,ЧызёЪиЯрЙиЗЈТЩ |
| ЦРТлМгдижа... |
| | ЩшЮЊЪзвГ | МгШыЪеВи | СЊЯЕеОГЄ | RSSЕМКНвГУц | АцШЈЩъУї |ЭјеОЕиЭМ | вНбЇТлЬГ | | |||||
|