



ЎЎЎЎ6Ј®Оҙ·Ц»ҜөДјдідЦКПё°ы Оҙ·Ц»ҜөДјдідЦКПё°ыЈЁundifferentiated mesenchymal cell)КЗұЈБфФЪіЙМеҪбөЮЧйЦҜДЪөДТ»Р©ҪПФӯКјөДПё°ыЈ¬ЛьГЗұЈіЦЧЕјдідЦКПё°ыөД·Ц»ҜЗұДЬЈ¬ФЪСЧЦўУлҙҙЙЛКұҝЙФцЦі·Ц»ҜОӘіЙПЛО¬Пё°ыЎўЦ¬·ҫПё°ыЎЈјдідЦКПё°ыіЈ·ЦІјФЪРЎСӘ№ЬУИЖдКЗГ«ПёСӘ№ЬЦЬО§Ј¬ІўДЬ·Ц»ҜОӘСӘ№ЬұЪөДЖҪ»¬јЎәНДЪЖӨПё°ыЎЈ
ЎЎЎЎ7Ј®°ЧПё°ыСӘТәДЪөД°ЧПё°ыЈ¬КЬЗч»ҜТтЧУөДОьТэЈ¬іЈҙ©іцГ«ПёСӘ№ЬәНОўҫІВцЈ¬УОЧЯөҪКиЛЙҪбөЮЧйЦҜДЪЈ¬РРК№Жд№ҰДЬЈ¬ІОУлГвТЯУҰҙрәНСЧЦў·ҙУҰЎЈКиЛЙҪбөЮЧйЦҜДЪТФКИЛбРФБЈПё°ыЎўБЬ°НПё°ыЎўЦРРФБЈПё°ы¶ајыЎЈУОЧЯіцөДөҘәЛПё°ыҪ«·Ц»ҜОӘҫЮКЙПё°ыЎЈ
ЎЎЎЎЈЁ¶ю)ПЛО¬
ЎЎЎЎ1Ј®ҪәФӯПЛО¬ ҪәФӯПЛО¬ЈЁcollagenous fiber)КэБҝЧо¶аЈ¬РВПККұіК°ЧЙ«Ј¬УР№вФуЈ¬УЦГы°ЧПЛО¬ЎЈHe ИҫЙ«ЗРЖ¬ЦРіККИЛбРФЈ¬ЧЕЗіәмЙ«ЎЈПЛО¬ҙЦПёІ»өИЈ¬Цұҫ¶1Јӯ20ҰМmЈ¬іКІЁАЛРОЈ¬Іў»ҘПаҪ»ЦҜЎЈҪәФӯФӯПЛО¬УЙЦұҫ¶20Ў«200nmөДҪәФӯФӯПЛО¬ХіәП¶шіЙЈЁНј3-2)ЎЈөзҫөПВЈ¬ҪәФӯФӯПЛО¬ПФГч°өҪ»МжөДЦЬЖЪРФәбОЖЈ¬әбОЖЦЬЖЪФј64nmЈЁНј3Јӯ12)ЎЈҪәФӯПЛО¬өДИНРФҙуЈ¬ҝ№АӯБҰЗҝЎЈҪәФӯПЛО¬өД»ҜС§іЙ·ЦОӘўсРНәНўтРНҪәФӯө°°ЧЎЈҪәФӯө°°ЧЈЁјтіЖҪәФӯЈ¬collagen)ЦчТӘУЙіЙПЛО¬Пё°ы·ЦГЪЎЈ·ЦГЪөҪПё°ыНвөДҪәФӯФЩҫЫәПіЙҪәФӯФӯПЛО¬Ј¬Ҫш¶шјҜәПіЙҪәФӯПЛО¬ЎЈ
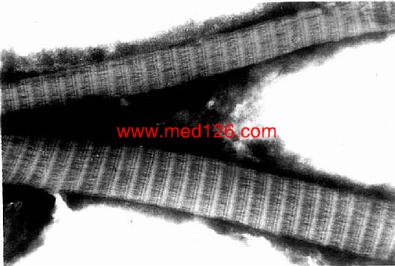
Нј3-12 ИЛҪәФӯПЛО¬өзҫөПсЎБ150000
ЈЁН¬јГТҪҝЖҙуС§ОдЦТецҪМКЪ№©Нј)
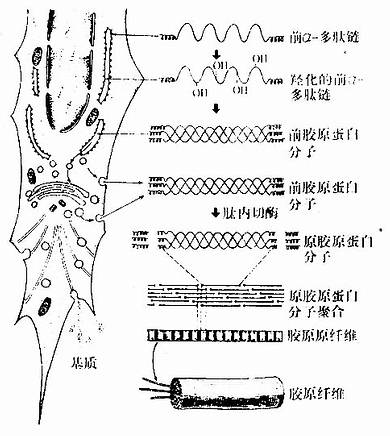
Нј3Јӯ13ҪәФӯПЛО¬Ул»щЦКРОіЙ№эіМКҫТвНј
ЎЎЎЎҪәФӯПЛО¬РОіЙөД»щұҫ№эіМИзПВЈЁНј3Јӯ13)Јә
ЎЎЎЎЈЁ1)Пё°ыДЪәПіЙЗ°ҪәФӯө°°Ч·ЦЧУЈәіЙПЛО¬Пё°ыЙгИЎәПіЙө°°ЧЦКЛщРиөД°ұ»щЛбЈ¬°ьАЁё¬°ұЛбЎўАө°ұЛбәНёК°ұЛбЈ¬ФЪҙЦГжДЪЦКНшөДәЛМЗМеЙП°ҙХХМШ¶ЁөДҪәФӯmRNAөДјо»щРтБРЈ¬әПіЙЗ°ҰБЈӯ¶алДБҙЎЈәуХЯұЯәПіЙұЯҪшИлҙЦГжДЪЦКНшЗ»ДЪЈ¬ІўФЪфЗ»ҜГёөДЧчУГПВЈ¬Ҫ«лДБҙЦРөДё¬°ұЛбәНАө°ұЛбфЗ»ҜЎЈҫӯфЗ»ҜәуЈ¬ИэМхЗ°ҰБЈӯ¶алДБҙ»ҘПаІшИЖіЙЙюЛчЧҙөДЗ°ҪәФӯө°°Ч·ЦЧУЈЁprocollagen molecule)ЎЈИЬҪвЧҙМ¬өДЗ°ҪәФӯө°°Ч·ЦЧУЈ¬БҪ¶ЛОҙІшИЖЈ¬іКЗтЧҙ№№РНЈ¬ФЪҙЦГжДЪЦКНшЗ»ДЪ»тЧӘТЖөҪёЯ¶ы»щёҙәПМеДЪјУИлМЗ»щәуЈ¬·ЦГЪөҪПё°ыНвЎЈ
ЎЎЎЎЈЁ2)ФӯҪәФӯө°°Ч·ЦЧУөДПё°ыНвҫЫәПЈәПё°ыНвөДЗ°ҪәФӯө°°Ч·ЦЧУЈ¬ФЪлДДЪЗРГёөДЧчУГПВЈ¬ЗРИҘ·ЦЧУБҪ¶ЛЗтЧҙ№№РОІҝ·ЦЈ¬РОіЙФӯҪәФӯө°°Ч·ЦЧУЈЁtropocol-lagen)ҙЦФј1.5nmЈ¬іӨФј300nmЎЈФӯҪәФӯө°°Ч·ЦЧУЖҪРРЕЕБРҫЫәПіЙҪәФӯФӯПЛО¬ЎЈҫЫәПКұЈ¬Па»ҘЖҪРРөДПаБЪ·ЦЧУҙнҝӘ1ЈҜ4·ЦЧУіӨ¶ИЈ¬Н¬Т»ЕЕөД·ЦЧУЈ¬КЧОІПа¶ФІўұЈіЦТ»¶ЁҫаАлЈ¬ҫЫәПіЙКшЈ¬УЪКЗРОіЙҫЯУР64nmЦЬЖЪәбОЖөДҪәФӯФӯПЛО¬ЎЈҫЫәПКұЈ¬·ЦЧУДЪЎў·ЦЧУјдөД»ҜС§»щТтҪшРРЛхәПЎўҪ»БӘЈ¬ФцјУФӯПЛО¬өДОИ№МРФЎЈИфёЙҪәФӯФӯПЛО¬ҫӯМЗө°°ЧХіәПіЙҙЦПёІ»өИөДҪәФӯПЛО¬ЎЈ
ЎЎЎЎҪәФӯПЛО¬өДТ»ІЛіЙКЬ¶а·ҪГжөДУ°ПмәНөчҝШЎЈИзПё°ыДЪё¬°ұЛбөДә¬БҝЦұҪУУ°ПмЗ°ҰБЈӯ¶алДБҙөДәПіЙЎЈИұСх»тИұ·ҰО¬ЙъЛШC»тFe2+өИёЁЦъТтЧУЈ¬өјЦВЗ°ҰБЈӯ¶алДБҙөДфЗ»ҜКЬөҪТЦЦЖЈ¬ФміЙЗ°ҪәФӯө°°ЧәПіЙХП°ӯЈ¬У°ПмҙҙЙЛөДУъәПЎЈҫЫәПКұЈ¬ИзҪәФӯө°°Ч·ЦЧУДЪәН·ЦЧУјдөДҪ»БӘХП°ӯЈЁіЈТтАө°ұхЈСх»ҜГёІ»ЧгЛщЦВ)Ҫ«У°ПмҪәФӯПЛО¬өДОИ№МРФЎЈіэіЙПЛО¬Пё°ыНвЈ¬іЙ№ЗПё°ыЎўИн№ЗПё°ыЎўДіР©ЖҪ»¬јЎПё°ыөИЖрФҙУЪјдідЦКөДПё°ыТФј°¶аЦЦЙПЖӨПё°ыТІДЬІъЙъҪәФӯө°°ЧЎЈ
ЎЎЎЎІ»Н¬ЧйЦҜөДҪәФӯө°°ЧЖд·ЦЧУАаРНІ»Н¬Ј¬ТСЦӨКөҰБЈӯ¶алДБҙ°ҙЖдТ»ј¶Ҫб№№·ЦОӘҰБ1Ј¬ҰБ2Ј¬ҰБ3Ј¬ИэАаЈ¬ёчАаУЦ·ЦОӘ10РНЈ¬ИзҰБ1ЈЁўс)ЎўҰБ1ЈЁўт)ЎўҰБ1ЈЁўу)ЎўҰБ1ЈЁўу)ЎӯЎӯҰБ1ЈЁX)ЎЈ
ЎЎЎЎёщҫЭ№№іЙҪәФӯө°°ЧИэ№ЙлДБҙөДІ»Н¬Ј¬ПЦТС·ўПЦУР11ЦЦІ»Н¬АаРНөДҪәФӯЎЈПЦҪ«ЦчТӘјёЦЦАаРНөДЧйіЙЎў·ЦІјәНМШөгБРҫЩУЪұнЈЁұн3Јӯ1)ЎЈ
ұн3Јӯ1 ҪәФӯө°°ЧөДАаРНЎў·ЦІјәНМШөг
| АаРН | З°ҪәФӯө°°ЧөДИэ№ЙлДБҙ | ·Ц Іј | Цч ТӘ МШ өг |
| ўс | [ҰБ1(ўс)]2ҰБ2(ўс) | ХжЖӨЎўҪоДӨЎў№®ДӨЎўұ»ДӨЎўлмЎўПЛО¬Ин№ЗЎў№ЗЎўСАұҫЦК | №№іЙЦВГЬІўУРәбОЖөДҙЦПЛО¬КшЈ¬ҝ№АӯБҰЗҝ |
| ўт | [ҰБ1(ўт)]3 | НёГчИн№ЗәНөҜРФИн№З | №№іЙУРәбОЖөДПёФӯПЛО¬Ј¬ҝ№С№БҰҪПЗҝ |
| ўу | [ҰБ1ЈЁўу)]3
[ҰБ1ЈЁўф)]2ҰБ2ЈЁўф) |
НшЧҙПЛО¬ЎўЖҪ»¬јЎЎўЙсҫӯДЪДӨЎў¶ҜВцЎўёОЎўЖўЎўЙцЎў·ОЎўЧУ№¬ | №№іЙУРәбОЖөДПёФӯПЛО¬Ј¬О¬іЦЖч№ЩөДРОМ¬Ҫб№№ |
| ўф | [ҰБ1ЈЁўф)]3
[ҰБ2ЈЁўф)]3 [ҰБ1ЈЁўх)]2ҰБ2ЈЁўх) |
»щДӨ»щ°еЎўҫ§ ЧҙМеДТ | І»РОіЙФӯПЛО¬Ј¬ОӘҫщЦКЧҙДӨЈ¬Ц§іЦәНВЛ№эЧчУГ |
| ўх | [ҰБ1ЈЁўх)]3
ҰБ1ЈЁўх)ҰБ2ЈЁўх)ҰБ3ЈЁўх) |
МҘДӨЎўјЎЎўлмЗК | №№іЙПёөДОЮәбОЖФӯПЛО¬ |
ЎЎЎЎ2Ј®өҜРФПЛО¬ өҜРФПЛО¬ЈЁelastic fiber)РВПКЧҙМ¬ПВіК»ЖЙ«Ј¬УЦГы»ЖПЛО¬ЎЈФЪHEұкұҫЦРЈ¬ЧЕЙ«ЗбОўЈ¬І»ТЧУлҪәФӯПЛО¬Зш·ЦЎЈө«И©ёҙәмЈЁaldehyde-fuchsin)»төШТВәмЈЁorcein)ДЬҪ«өҜРФПЛО¬ИҫіЙЧПЙ«»тЧШәЦЙ«ЎЈөҜРФПЛО¬ҪППёЈ¬ЦұРР,·ЦЦ§Ҫ»ЦҜЈ¬ҙЦПёІ»өИЈЁ0.2Јӯ1.0ҰМm),ұнГж№в»¬Ј¬¶П¶ЛіЈҫнЗъЈЁНј3Јӯ2)ЎЈөзҫөПВЈ¬өҜРФПЛО¬өДәЛРДІҝ·ЦөзЧУГЬ¶ИөНЈ¬УЙҫщЦКөДөҜРФө°°ЧЈЁelastin)ЧйіЙЈ¬әЛРДНвЦЬёІёЗОўФӯПЛО¬ЈЁmicrofibril)Ј¬Цұҫ¶Фј10nmЎЈөҜРФө°°Ч·ЦЧУДЬИОТвҫнЗъЈ¬·ЦЧУјдҪе№ІјЫјьҪ»БӘіЙНшЎЈФЪНвБҰЗЈАӯПВЈ¬ҫнЗъөДөҜРФө°°Ч·ЦЧУЙмХ№АӯіӨЈ»іэИҘНвБҰәуЈ¬өҜРФө°°Ч·ЦЧУУЦ»ШёҙОӘҫнЗъЧҙМ¬ЈЁНј3Јӯ14)ЎЈ
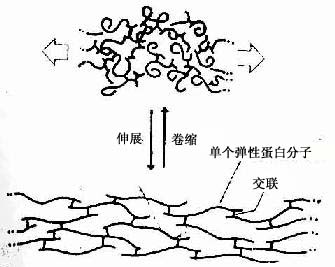
Нј3-14 ЙмЛхЧҙМ¬ПВөҜРФө°°Ч·ЦЧУөД№№РОұд»Ҝ
ЎЎЎЎөҜРФПЛО¬ё»УЪөҜРФ¶шИНРФІоЈ¬УлҪәФӯПЛО¬Ҫ»ЦҜФЪТ»ЖрЈ¬К№КиЛЙҪбөЮЧйЦҜјИУРөҜРФУЦУРИНРФЈ¬УРАыУЪЖч№ЩәНЧйЦҜұЈіЦРОМ¬О»ЦГөДПа¶Фәг¶ЁЈ¬УЦҫЯУРТ»¶ЁөДҝЙұдРФЎЈ
ЎЎЎЎ3Ј®НшЧҙПЛО¬ НшЧҙПЛО¬ЈЁreticular fiber)ҪППёЈ¬·ЦЦ§¶аЈ¬Ҫ»ЦҜіЙНшЎЈНшЧҙПЛО¬УЙўуРНҪәФӯө°°Ч№№іЙЈ¬ТІҫЯУР64nmЦЬЖЪРФәбОЖЎЈПЛО¬ұнГжұ»ёІө°°Ч¶аМЗәНМЗө°°ЧЈ¬№КPAS·ҙУҰСфРФЈ¬ІўҫЯКИТшРФЎЈУГТшИҫ·ЁЈ¬НшЧҙПЛО¬іКәЪЙ«Ј¬№КУЦіЖКИТшПЛО¬ЈЁargyrophil fiber)ЎЈНшЧҙПЛО¬¶а·ЦІјФЪҪбөЮЧйЦҜУлЖдЛьЧйЦҜҪ»ҪзҙҰЈ¬Из»щДӨөДНш°еЎўЙцРЎ№ЬЦЬО§ЎўГ«ПёСӘ№ЬЦЬО§ЎЈФЪФмСӘЖч№ЩәНДЪ·ЦГЪПЩЈ¬УРҪП¶аөДНшЧҙПЛО¬Ј¬№№іЙЛьГЗөДЦ§јЬЎЈ
ЎЎЎЎЈЁИэ)»щЦК
ЎЎЎЎ»щЦКЈЁground substance)КЗТ»ЦЦУЙЙъОпҙу·ЦЧУ№№іЙөДҪәЧҙОпЦКЈ¬ҫЯУРТ»¶ЁХіРФЎЈ№№іЙ»щЦКөДҙу·ЦЧУОпЦК°ьАЁө°°Ч¶аМЗәНМЗө°°ЧЎЈ
ЎЎЎЎө°°Ч¶аМЗЈЁproteoglycan)КЗУЙө°°ЧЦКУлҙуБҝ¶аМЗҪбәПіЙөДҙу·ЦЧУёҙәПОпЈ¬КЗ»щЦКөДЦчТӘіЙ·ЦЎЈЖдЦР¶аМЗЦчТӘКЗНёГчЦКЛбЈЁhyaluronic acid)Ј¬ЖдҙОКЗБтЛбИн№ЗЛШA ЎўCЈЁchondroitin sulfate AЎўC)ЎўБтЛбҪЗЦКЛШAЎўCЈЁkeratin sulfate)БтЛбТТхЈёОЛШЈЁheparan sulfate)өИЎЈЛьГЗ¶јКЗТФә¬УР°ұ»щТСМЗөДЛ«МЗОӘ»щұҫөҘО»ҫЫәПіЙөДіӨБҙ»ҜәПОпЈ¬ЧЬіЖОӘМЗ°·¶аМЗЈЁglycosaminoglycan,GAG)ЎЈУЙУЪМЗ°·¶аМЗ·ЦЧУҙжФЪҙуБҝТхАлЧУЈ¬№КДЬҪбәПҙуБҝЛ®ЈЁҪбәПЛ®)ЎЈНёГчЦКЛбКЗТ»ЦЦЗъХЫЕМИЖөДіӨБҙҙу·ЦЧУЈ¬АӯЦұҝЙҙп2.5ҰМmЈ¬УЙЛь№№іЙө°°Ч¶аМЗёҙәПОпөДЦчёЙЈ¬ЖдЛьМЗ°·¶аМЗФтТФө°°ЧЦКОӘәЛРД№№іЙө°°Ч¶аМЗСЗөҘО»Ј¬әуХЯФЩНЁ№эБ¬ҪУө°°ЧҪбәПФЪНёГчЦКЛбіӨБҙ·ЦЧУЙПЈЁНј3-15)ЎЈө°°Ч¶аМЗёҙәПОпөДБўМе№№РНРОіЙУРРн¶аОўҝЧП¶өД·ЦЧУЙёЈ¬РЎУЪҝЧП¶өДЛ®әНИЬУЪЛ®өДУӘСшОпЎўҙъР»ІъОпЎўјӨЛШЎўЖшМе·ЦЧУөИҝЙТФНЁ№эЈ¬ұгУЪСӘТәУлПё°ыЦ®јдҪшРРОпЦКҪ»»»ЎЈҙуУЪҝЧП¶өДҙу·ЦЧУОпЦКЈ¬ИзПёҫъөИІ»ДЬНЁ№эЈ¬К№»щЦКіЙОӘПЮЦЖПёҫъА©ЙўөД·АУщЖБХПЎЈИЬСӘРФБҙЗтҫъәН°©Пё°ыөИДЬІъЙъНёГчЦКЛбГёЈ¬ЖЖ»ө»щЦКөД·АУщЖБХПЈ¬ЦВК№ёРИҫәНЦЧБцҪюИуА©ЙўЎЈ

Нј3-15 ө°°Ч¶аМЗ·ЦЧУҪб№№ДЈРН
ЎЎЎЎМЗө°°ЧЈЁglycoprotein)КЗ»щЦКДЪБнТ»АаЦШТӘөДЙъОпҙу·ЦЧУЈ¬Улө°°Ч¶аМЗПа·ҙЈ¬ЖдЦчТӘіЙ·ЦКЗө°°ЧЦКЎЈҙУ»щЦКДЪТСҫӯ·ЦАліц¶аЦЦМЗө°°ЧЈ¬ЦчТӘөДУРПЛО¬ХіБ¬ө°°ЧЈЁfibronectin FN)ІгХіБ¬ө°°ЧЈЁlaminin)әНИн№ЗХіБ¬ө°°ЧЈЁchondronectin)өИЎЈХвАа»щЦКҙу·ЦЧУІ»ҪцІОУл»щЦК·ЦЧУЙёөД№№іЙЈ¬Н¬КұНЁ№эЛьГЗөДБ¬ҪУәНҪйөјЧчУГТІУ°ПмПё°ыөДёҪЧЕәНТЖ¶ҜТФј°ІОУлөчҪЪПё°ыөДЙъіӨәН·Ц»ҜЎЈ
ЎЎЎЎПЛО¬ХіБ¬ө°°ЧКЗ»щЦКЦРТ»ЦЦЦШТӘөДМЗө°°ЧЈ¬ҙжФЪУЪҪәФӯПЛО¬әНРн¶аҪбөЮЧйЦҜПё°ыЦЬО§ЎЈФЪөзҫөПВЈ¬ПЛО¬ХіБ¬ө°°ЧіКФӯПЛО¬ЧҙЈ¬УЙБҪМх¶алДБҙЧйіЙЈ¬БҪМхлДБҙөДТ»¶ЛУЙИфёЙ¶юБтјьБ¬ҪУЎЈГҝТ»лДБҙЙПҫщУРИфёЙМШ¶ЁөД№ҰДЬЗшЈ¬ДЬ·ЦұрУлПё°ыЎўҪәФӯЎўёОЛШәНПЛО¬ЛШөИҪбәПЎЈУЪКЗЈ¬ПЛО¬ХіБ¬ө°°ЧЧчОӘТ»ЦЦЦРҪйө°°ЧЈ¬ДЬҪ«Пё°ыБ¬ҪУөҪҪәФӯЎўёОЛШөИПё°ыНв»щЦКЙПЎЈ
ЎЎЎЎЧйЦҜТәЈЁtissue fluid)КЗҙУГ«ПёСӘ№Ь¶ҜВц¶ЛЙшИл»щЦКДЪөДТәМеЈ¬ҫӯГ«ПёСӘ№ЬҫІВц¶ЛәНГ«ПёБЬ°Н№Ь»ШБчИлСӘТә»тБЬ°НЈ¬ЧйЦҜТәІ»¶ПёьРВЈ¬УРАыУЪСӘТәУлПё°ыҪшРРОпЦКҪ»»»Ј¬іЙОӘЧйЦҜәНПё°ыАөТФЙъҙжөДДЪ»·ҫіЎЈөұЧйЦҜТәөДЙшіцЎў»ШБч»т»ъМеЛ®СОЎўө°°ЧЦКҙъР»·ўЙъХП°ӯКұЈ¬»щЦКЦРөДЧйЦҜТәә¬БҝҝЙФц¶а»тјхЙЩЈ¬өјЦВЧйЦҜЛ®ЦЧ»тНСЛ®ЎЈ




