



一、心肌能量代谢障碍
(一)能量生成(释放)障碍
心肌主要借各种能源物质包括脂肪酸,葡萄糖等的有氧氧化而获得能量。心肌细胞对氧的需要量很大,摄取能力很强,在正常安静情况下,冠状动静脉血氧含量差可高达14ml%。可见,心肌氧供给不足或有氧氧化过程的障碍,均可使心肌细胞内能量生成不足而导致心肌收缩性减弱。
严重的贫血、冠状动脉硬化等所引起的心肌缺氧,是导致心肌细胞内能量生成不足的常见原因。维生素B1缺乏时,由于焦磷酸硫胺素(丙酮酸脱羧酶的辅酶)生成不足,丙酮酸的氧化发生障碍,故也可引起心肌能量生成不足。肥大的心肌也可因心肌缺氧而导致能量生成不足(详后文)。医学全在线www.med126.com
(二)能量利用障碍
心肌细胞内氧化磷酸化过程中所产生的ATP,在心肌兴奋-收缩偶联过程中受到肌球蛋白头部ATP酶的作用而水解,为心肌收缩提供能量。实验表明,部分动物的心肌由肥大转向衰竭时,心肌耗氧量和ATP含量并不减少而完成的机械功却显著减少,说明心肌利用ATP中的化学能作机械功的过程有障碍,即心肌的能量利用发生障碍。有人发现,随着心肌负荷过重而发生心肌肥大时,心肌收缩蛋白的结构发生变化,肌球蛋白头部ATP酶的活性降低,ATP水解发生障碍,因此能量利用发生障碍,心肌收缩性乃因而减弱。这种现象也可见于老年人及甲状腺功能低下的心脏。关于心肌收缩蛋白质结构发生变化的机制尚未阐明。
二、兴奋-收缩偶联障碍-Ca2+的运转失常
近年来,在心力衰竭的发病机制中,因Ca2+运转失常引起的心肌兴奋-收缩偶联障碍,受到了很大重视。正常心肌在复极化时,心肌细胞内肌质网的ATP酶(钙泵)被激活,从而使胞质中的Ca2+逆着浓度差被摄取到肌质网中储存;同时,另一部分Ca2+则从胞质中被转运到细胞外。于是心肌细胞胞质Ca2+浓度降低。心肌舒张。心肌除极化时,肌质网向胞质释放Ca2+,同时又有Ca2+从细胞外液进入胞质,因而胞质中Ca2+浓度增高,心肌收缩。心肌兴奋-收缩偶联障碍的发生机制主要有:
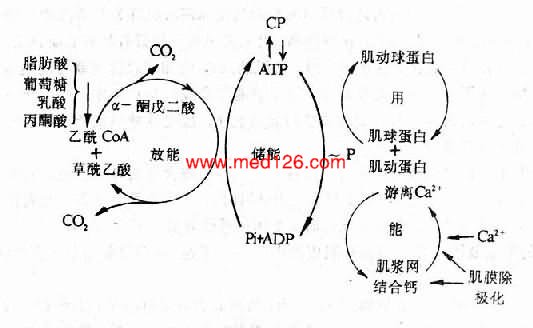
图12-3 心肌能量代谢
(一)肌质网摄取Ca2+减少有人发现,在过度肥大的心肌中,肌质网ATP酶的活性降低,因而在心肌复极化时肌质网摄取和储存Ca2+的量减少,除极化时肌质网向胞质释放的Ca2+也因之减少。由此所引起的心肌细胞除极化时胞质内Ca2+浓度的低下可能是心肌收缩性减弱的重要原因。
另据报道,在肌质网摄取Ca2+减少的同时,线粒体对Ca2+摄取量增多,但线粒体在心肌除极化时向胞质释放Ca2+的速度却非常缓慢。Ca2+在心肌细胞中这种异常的分布也是胞质Ca2+浓度降低的一个原因。此外,还有人认为线粒体内Ca2+的增多可引起氧化磷酸化脱偶联,从而使能量生成不足。
(二)酸中毒和高钾血症 Ca2+的运转也受H+和K+的影响。在心力衰竭时有一定程度的缺氧,故可有细胞外液H+和K+浓度的增高。关于H+如何影响运转的问题也未完全清楚。不同的作者提出过不同的假设:Katz等曾认为H+能在肌钙蛋白上与Ca2+竞争结合位置,因而在H+过多时就能取代Ca2+的位置而使心肌的兴奋-收缩偶联发生障碍。也有人报道,在H+浓度增高时,Ca2+与肌质网的结合比较牢固,除极化时肌质网释放Ca2+减少,故H+增多时,心肌的兴奋-收缩偶联发生障碍。前文已经提到,正常心肌细胞在除极化时有Ca2+从细胞外液进入胞质,而最近有人报道,在细胞外液H+浓度增高时,Ca2+的内流减慢,故心肌兴奋-收缩偶联发生障碍。
细胞外液中的K+和Ca2+在心肌细胞上有互相竞争的作用。当外液中K+浓度升高时,动作电位中Ca2+内流就减少,因而心肌胞质中Ca2+浓度降低,这也是引起心肌兴奋-收缩偶联障碍的一个因素。




