



随着DNA polⅢ向前移动,先导链的合成逐渐延长的同时,岗崎片段也在不断延长,这一噜噗也在不断扩大。当岗崎片段合成到前一个片段的5′端时,这一大噜噗就释放出来,由于复制叉向前移动又可将另一部分随从链的模板置换出来,由引发体合成新的引物,然后再形成一个小的噜噗,进行新的岗崎片段的合成。由此模型不难看出:随从链的合成需要周期性的引发,因此其合成进度总是与前导链相差一个岗崎片段的长度。岗崎片段完成后,其5′端的RNA引物由DNa polⅠ的5′→3′外切酶活性切除,由此造成的空隙再由DNA polⅠ的5′→3′聚合活性催化dNTP得到填补。所以DNA的复制是在DNa polⅢ和DNA polⅠ互相配合下完成的。
下面列表说明三种大肠杆菌DNA聚合酶的特性
表16-1 大肠杆菌DNA聚合酶特征
| DNA聚合酶Ⅰ | DNA聚合酶Ⅱ | DNA聚合酶Ⅲ | |
| 分子量 | 109KD | 120KD | >600KD |
| 每个细胞中的分子数 | 400 | 17-100 | 10-20 |
| 5′→3′聚合活性 | + | + | + |
| 37℃转化率核苷酸数/酶分子·分钟 | 600 | 30 | 30,000 |
| 5′→3′外切活性 | + | - | - |
| 5′→3′外切活性 | + | + | + |
| 切刻平移活性 | + | - | - |
| 对dNTP亲和力 | 低 | 低 | 高 |
| 功能 | 修复 | 不详 | 复制 |
| 去除引物 | |||
| 填补空缺 |
真核生物DNA聚合酶
真核生物DNA聚合酶有α、β、γ、δ及ε。它们的基本特性相似于大肠杆菌DNA聚合酶,其主要活性是催化dNTP的5′→3′聚合活性,基本特征见表16-2。
表16-2 真核生物DNA聚合酶
| α | β | γ | δ | ε | |
| 亚基数 | 4 | 4 | 4 | 2 | 5 |
| 分子量(KD) | >250 | 36-38 | 160-300 | 170 | 256 |
| 细胞内定位 | 核 | 核 | 线粒体 | 核 | 核 |
| 5′→3′聚合活性 | + | + | + | + | + |
| 3′→5′外切活性 | - | - | - | - | - |
| 功能 | 复制、引发 | 修复 | 复制 | 复制 | 复制 |
真核细胞在DNA复制中起主要作用的是DNA polα,主要负责染色体DNA的复制。DNa polβ的模板特异性是具有缺口的DNA分子,被认为它与DNA修复有关。DNa polγ在线粒体DNA的复制中起作用。DNA polδ不但有5′→3′聚合活性,而且还具有3′→5′外切酶活性,据认为真核生物DNA复制是在DNa polα和DNA polδ协同作用下进行的,前导链的合成靠DNA polδ催化,并且还需要一种细胞周期调节因子椩鲋诚赴丝乖?proliferating cell nucleus antigen, PCNA)参与。而随从链的合成靠DNA polα和引发酶配合作用完成。
(二)与超螺旋松驰有关的酶:
DNA复制从起始点开始向一个方向复制时,局部的DNA双链必须打开,主要靠解链酶的作用,打开后的单链还需要单链结合蛋白与其结合,在复制叉向前移动时造成其前方DNA分子所产生的正超螺旋,必须由拓扑异构酶来解决。下面分别介绍它们的作用。
拓扑异构酶(topoisomerase)是一类改变DNA拓扑性质的酶。在体外可催化DNA的各种拓扑异构化反应,而在生物体内它们可能参与了DNA的复制与转录。在DNA复制时,复制叉行进的前方DNA分子部分产生有正超螺旋,拓扑酶可松驰超螺旋,有利于复制叉的前进及DNA的合成。DNA复制完成后,拓扑酶又可将DNA分子引入超螺旋,使DNA缠绕、折叠,压缩以形成染色质。DNA拓扑异构酶有Ⅰ型和Ⅱ型,它们广泛存在于原核生物及真核生物中。表16-3
表16-3 大肠杆菌和真核生物中的拓扑异构酶
| 类型 | 作用 | 对超螺旋的作用 |
| Ⅰ型拓扑异构酶 | ||
| 大肠杆菌 | 切开一股DNA链 | 松驰负超螺旋 |
| 真核生物 | 切开一股DNA链 | 松驰正,负超螺旋 |
| Ⅱ型拓扑异构酶 | ||
| 大肠杆菌 | 切开二股DNA链 | 构驰正超螺旋; |
| 依赖ATP | 引入负超螺旋, | |
| 解环连等 | ||
| 真核生物 | 切开二股DNA链 | 松驰正超旋, |
| 依赖ATP | 但不能引入负超螺旋 |
拓扑异构酶Ⅰ(TopoⅠ)的主要作用是将环状双链DNA的一条链切开一个口,切口处链的末端绕螺旋轴按照松驰超螺旋的方向转动,然后再将切口封起来。这就使DNA复制叉移动时所引起的前方DNA正超螺旋得到缓解,利于DNA复制叉继续向前打开。拓扑异构酶Ⅰ除上述作用外,对环状单链DNA还有打结或解结作用,对环状双链DNA的环连或解连以及使环状单链DNA形成环状双链DNA都有作用(图16-13)。
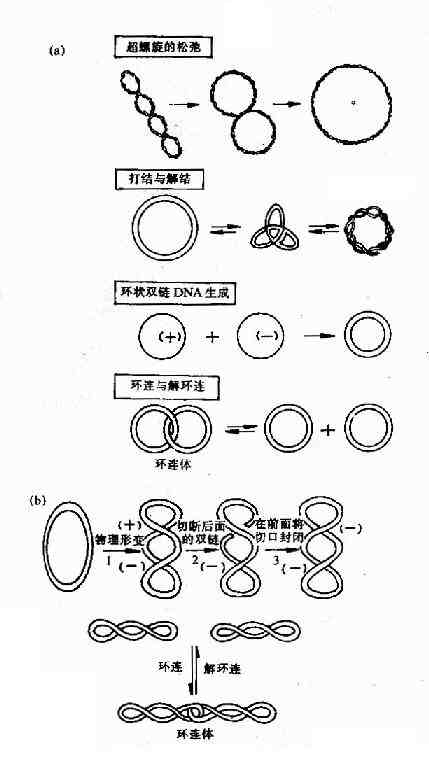
图16-13 拓扑酶Ⅰ及Ⅱ的作用特点
(a)大肠杆菌拓扑酶Ⅰ催化的4种拓扑异构化作用 (b)拓扑酶Ⅱ的作用
拓扑异构酶Ⅱ(TopoⅡ)是在大肠杆菌中发现的,曾被称为旋转酶(gyrase),它们作用特点是切开环状双链DNA的两条链,分子中的部分经切口穿过而旋转,然后封闭切口,TopoⅡ还可使DNA分子从超螺旋状态转变为松驰状态,此反应不需要ATP参与。DNA复制完成后,TopoⅡ在ATP参与下,DNA分子从松驰状态转变为负超螺旋。此外,TopoⅡ催化的拓扑异构化反应还有环连或解环连,以及打结或解结。




