



四、DNA复制的终止阶段
DNA在复制过程中,合成出的前导链为一条连续的长链。随从链则是由合成出许多相邻的片段,在连接酶的催化下,连接成为一条长链。连接作用是在连接酶催化下进行的。
连接酶(ligase)的作用是催化相邻的DNA片段以3′、5′-磷酸二酯键相连接。连接反应中的能量来自ATP(或NAD+)。连接酶先与ATP作用,以共价键相连生成E桝MP中间体。中间体即与一个DNA片段的5′-磷酸相连接形成E-AMP-5′-DNA。然后再与另一个DNA片段的3′-OHH末端作用,E和AMP脱下,两个DNA片段以3′、5′磷酸二酯键相连接。随从链的各个DNA片段就是这样连接成一条DNA长链(图16-14)。
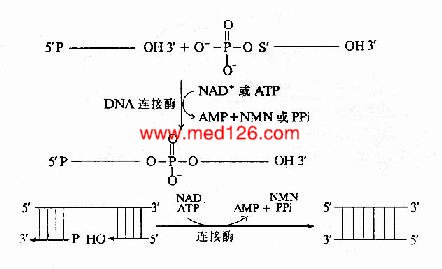
图16-14 连接酶的催化反应
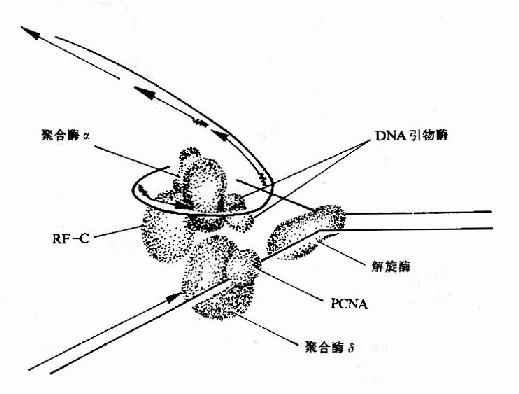
图16-15 真核生物DNA复制叉结构示意图
已有研究证明大肠杆菌染色体DNA具有复制终止位点,此处可以结合一种特异的蛋白质分子叫做Tus,这个蛋白质可能是通过阻止解链酶(Helicase)的解链活性而终止复制的。详细的机制还不完全清楚。
DNA复制完成后,靠拓扑酶将DNA分子引入超螺旋结构。
五、真核生物DNA复制的特点:
DNA复制的研究最初是在原核生物中进行的,有些原核生物的DNA复制已经搞得很清楚。真核生物比原核生物复杂得多,但DNA复制的基本过程还是相似的。在这里我们主要讨论一些重要的区别。
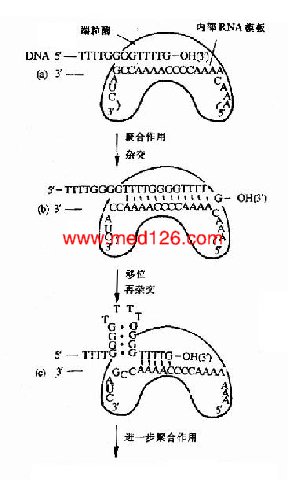
图16-16 端粒酶催化端区TG链的合成
1.与原核生物不同,真核生物DNA复制有许多起始点,例如酵母S.cerevisiae的17号染色体约有400个起始点,因此,虽然真核生物DNA复制的速度(60核苷酸/每秒钟)比原核生物DNA复制的速度(E.coli 1700核苷酸/每秒钟)慢得多,但复制完全部基因组DNA也只要几分钟的时间。
2.SV40病毒DNA主要依靠宿主细胞中的DNA复制体系进行DNA的复制,这是了解真核生物DNA复制的体外模型。在真核生物DNA复制叉处,需要两种不同的酶。DNA聚合酶α(polα)和DNA聚合酶δ(polδ)。polα和引物酶紧密结合,在DNA模板上先合成RNA引物,再由polα延长DNA链,这种活性还要复制因子C参与。同时结合在引物模板上的PCNA(增殖细胞核抗原Proliferating cell nuclear antigen)此时释放了polα,然后由polδ结合到生长链3′末端,并与PCNA结合,继续合成前导链。而随从链的合成靠polα紧密与引物酶结合并在复制因子C帮助下,合成岗崎片段(图16-15)。
3.由于真核生物染色体是线性DNA,它的两端叫做端区(telomeres),端区是由重复的寡核苷酸序列构成的。例如酵母的端区重复序列是5′G(1?)T(3)3′。前面讲到所有生物DNA聚合酶都只能催化DNA从5′→3′的方向合成,因此当复制叉到达线性染色体末端时,前导链可以连续合成到头,而由于随从链是以一种不连续的形式合成岗崎片段,所以不能完成线性染色体末端的复制,如果这个问题不解决,真核生物在细胞分裂时DNA复制将产生5′末端隐缩,使DNA缩短,近十多年的研究表明,真核生物体内都存在一种特殊的反转录酶叫做端粒酶(telomerase),它是由蛋白质和RNA两部分组成的,它以自身的RNA为模板,在随从链模板DNA的3′桹H末端延长DNA,再以这种延长的DNA为模板,继续合成随从链(图16-16)。
由此可见端粒酶在保证染色体复制的完整性上有重要意义。




