



ЎЎЎЎ1979ДкЈ¬WangәНRichөИИЛФЪСРҫҝИЛ№ӨәПіЙөДCGCGCGөҘҫ§өДX-ЙдПЯСЬЙдНјЖЧКұіцИЛТвБПөШ·ўПЦХвЦЦБщҫЫМеөД№№ПуУлЙПГжҪІөҪөДНкИ«І»Н¬ЎЈЛьКЗЧуКЦЛ«ВЭРэЈ¬УлУТКЦВЭРэөДІ»Н¬КЗВЭҫаСУіӨ(4.5nmЧуУТ)Ј¬Цұҫ¶ұдХӯ(1.8nm)Ј¬ГҝёцВЭРэә¬12ёцјо»щ¶ФЈ¬·ЦЧУіӨБҙЦРБЧФӯЧУІ»КЗЖҪ»¬СУЙм¶шКЗҫвіЭРОЕЕБРЈ¬УРИзЎ°Ц®ЎұЧЦРОТ»СщЈ¬ТтҙЛҪРЛьZ№№Пу(УўОДЧЦZigzagөДөЪТ»ёцЧЦДё)ЎЈ»№УРЈ¬ХвТ»№№ПуЦРөДЦШёҙөҘО»КЗ¶юәЛЬХЛб¶шІ»КЗөҘәЛЬХЛбЈ»¶шЗТZӘІDNAЦ»УРТ»ёцВЭРэ№өЈ¬ЛьПаөұУЪB№№ПуЦРөДРЎ№өЈ¬ЛьПБ¶шЙоЈ¬ҙу№өФтІ»ёҙҙжФЪ(Нј15-7)ЎЈҪшТ»ІҪөД·ЦОц»№ЦӨГчЈ¬ZЈӯDNAөДРОіЙКЗDNAөҘБҙЙПіцПЦаСЯКУлаЧаӨҪ»МжЕЕБРЛщіЙөДЎЈұИИзCGCGCGCG»тХЯCACACACAЎЈ
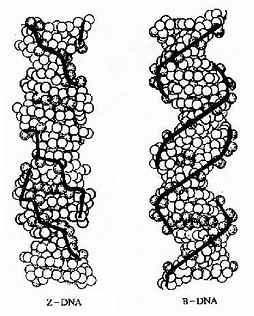
Нј15Јӯ7ЎЎZЈӯDNAәНBЈӯDNA
ЎЎЎЎZЈӯDNAУРКІГҙЙъОпС§ТвТеДШ?УҰөұЦёіцZЈӯDNAөДРОіЙНЁіЈФЪИИБҰС§ЙПКЗІ»АыөДЎЈТтОӘZЈӯDNAЦРҙшёәөзәЙөДБЧЛбёщҫаАлМ«ҪьБЛЈ¬Хв»бІъЙъҫІөзЕЕівЎЈө«КЗЈ¬DNAБҙөДҫЦІҝІ»ОИ¶ЁЗшөДҙжФЪҫНіЙОӘЗұФЪөДҪвБҙО»өгЎЈDNAҪвВЭРэИҙКЗDNAёҙЦЖәНЧӘВјөИ№эіМЦРұШТӘөД»·ҪЪЈ¬ТтҙЛИПОӘХвТ»Ҫб№№Ул»щТтөчҪЪУР№ШЎЈұИИзSV40ФцЗҝЧУЗшЦРҫНУРҙЛҪб№№Ј¬УЦИзКуАаОўРЎІЎ¶ҫDNSёҙЦЖЗшЖрКјөгёҪҪьУРGCҪ»МжЕЕБРРтБРЎЈҙЛНвЈ¬DNAВЭРэЙП№өөДМШХчФЪЖдРЕПўұнҙп№эіМЦРЖр№ШјьЧчУГЎЈөчҝШө°°Ч¶јКЗНЁ№эЖд·ЦЧУЙПМШ¶ЁөД°ұ»щЛбІаБҙУлDNAЛ«ВЭРэ№өЦРөДјо»щ¶ФТ»ІаөДЗвФӯЧУ№©Ме»тКЬМеПа»ҘЧчУГЈ¬РОіЙЗвјьҙУ¶шК¶ұрDNAЙПөДТЕҙ«РЕПўөДЎЈҙу№өЛщҙшөДТЕҙ«РЕПўұИРЎ№ө¶аЎЈ№өөДҝнХӯәНЙоЗіТІЦұҪУУ°ПмөҪөчҝШө°°ЧЦК¶ФDNAРЕПўөДК¶ұрЎЈZӘІDNAЦРҙу№өПыК§Ј¬РЎ№өПБ¶шЙоЈ¬К№өчҝШө°°ЧК¶ұр·ҪКҪТІ·ўЙъұд»ҜЎЈХвР©¶ј°өКҫZӘІDNAөДҙжФЪІ»ҪцҪцКЗУЙУЪDNAЦРіцПЦаСЯКТ»аӨаЧҪ»МжЕЕБРЦ®Ҫб№ыЈ¬ТІТ»¶ЁКЗФЪВюВюөДҪш»ҜіӨәУЦР¶ФDNAРтБРУлҪб№№І»¶ПөчХыУлЙёСЎөДҪб№ыЈ¬УРЖдДЪФЪ¶шЙоҝМөДә¬ТвЈ¬Ц»КЗИЛГЗ»№Оҙід·ЦИПК¶¶шТСЎЈ
ЎЎЎЎDNA№№ПуөДҝЙұдРФЈ¬»тХЯЛөDNA¶юј¶Ҫб№№өД¶аМ¬РФөД·ўПЦНШҝнБЛИЛГЗөДКУТ°ЎЈФӯАҙЈ¬ЙъОпМеЦРЧоОӘОИ¶ЁөДТЕҙ«ОпЦКТІҝЙТФІЙУГІ»Н¬өДЧЛМ¬АҙКөПЦЖд·бё»¶аІЙөДЙъОпС§№ҰДЬЎЈ
ЎЎЎЎ¶аДкАҙЈ¬DNAҪб№№өДСРҫҝКЦ¶ОЦчТӘКЗXЙдПЯСЬЙдјјКхЈ¬ЖдҪб№ыКЗНЁ№эјдҪУ№ЫІв¶аёцDNA·ЦЧУУР№ШҪб№№ІОКэөДЖҪҫщЦө¶ш»сөГөДЎЈН¬КұЈ¬ХвПојјКхөДСщЖ··ЦОцМхјюК№ұ»ІвDNA·ЦЧУУлМмИ»ЧҙМ¬ПаІоЙхФ¶ЎЈТтҙЛЈ¬ФЪ·ҙУіDNAҪб№№ХжКөРФ·ҪГжХвЦЦ·Ҫ·ЁҙжФЪЧЕИұПЭЎЈ1989ДкЈ¬УҰУГЙЁГиЛнөАПФОўҫө(scanning tummeling microscopy,STM)СРҫҝDNAҪб№№ҝЛ·юБЛЙПКцјјКхөДИұПЭЎЈХвЦЦПИҪшөДПФОўјјКхЈ¬І»ҪцҝЙҪ«ұ»ІвОп·Еҙу500Нтұ¶Ј¬ЗТДЬЦұҪУ№ЫІвҪУҪьМмИ»МхјюПВөҘёцDNA·ЦЧУөДҪб№№ПёҪЪЎЈSTMјјКхөДУҰУГКЗDNAҪб№№СРҫҝЦРөДЦШТӘҪшХ№Ј¬ҝЙНыФЪМҪЛчDNAҪб№№өДДіР©ОҙЦӘөгЙПХ№КҫҫЮҙуЗұБҰЎЈ
ЎЎЎЎ(Иэ)DNAҪб№№өДІ»ҫщТ»РФ(heterogeneity)
ЎЎЎЎФЪDNAөДТ»ј¶Ҫб№№ЦРЈ¬ЛДЦЦјо»щAЈ¬TЈ¬CЈ¬GФ¶·ЗҫщФИ·ЦІјЈ¬ҫЎ№ЬЛ«ВЭРэөД№№РНҙуМеПаН¬Ј¬ө«СШЧЕDNAБҙёчҙҰөДОпАнҪб№№І»НкИ«ПаН¬Ј¬ёчҙҰЛ«ВЭРэөДОИ¶ЁРФТІҫНПФКҫіцІоұрЈ¬ід·ЦМеПЦБЛDNAТ»ј¶Ҫб№№ҫц¶ЁёЯј¶Ҫб№№өДФӯАнЎЈЖдІ»ҫщТ»РФЦчТӘУРЈә
ЎЎЎЎ1.·ҙПтЦШёҙРтБР(inverted repeats)
ЎЎЎЎУЦіЖ»ШОДРтБР(palindrome)Ј¬ЛьДЬФЪDNA»тRNAЦРРОіЙ·ўјРҪб№№ЎЈХвЦЦ»ШОДҪб№№НЁіЈКЗЧчОӘТ»ЦЦМШұрРЕәЕЈ¬ИзПЮЦЖРФәЛЛбДЪЗРЛб(restriction enclгЕеДonuclease)ј°өчҪЪө°°ЧөДК¶ұрО»өгЈ¬ЧӘВјЦХЦ№РЕәЕөИЎЈ
ЎЎЎЎ2.ё»ә¬A/TөДРтБР
ЎЎЎЎФЪёЯөИЙъОпЦРЈ¬A+TУлGЈ«CөДә¬БҝІоІ»¶аПаөИЈ¬И»¶шФЪЛьГЗөДИҫЙ«МеДіТ»ЗшУтЈ¬AЎӨTә¬БҝҝЙДЬПаөұёЯЎЈИзФЪәЬ¶аУРЦШТӘөчҪЪ№ҰДЬөДDNAЗш¶О¶јё»ә¬AЎӨTЈ¬МШұрКЗФЪёҙЦЖЖрөгәНЖф¶ҜЧУөДPribnowҝт(ХжәЛЙъОпОӘTATAҝт)өДРтБРЦРЈ¬Жд¶ФУЪёҙЦЖәНЖрКјК®·ЦЦШТӘЎЈТтОӘAЈӯT¶ФЦ»УР¶юМхЗвјьЈ¬ҙЛҙҰөДЛ«БҙҪПGЈӯC¶ФҙҰТЧУЪҪвҝӘЈ¬УРАыУЪЖрКјёҙәПОпөДРОіЙЎЈ
ЎЎЎЎ3.аСЯКәНаЧаӨөДЕЕБРЛіРт¶ФЛ«ВЭРэҪб№№ОИ¶ЁРФөДУ°ПмЎЈ
ЎЎЎЎИЛГЗҝјІмБЛК®ЦЦПаБЪөД¶юәЛЬХЛб¶Ф(nearestӘІneighbor doublets)Ј¬·ўПЦТ»ёц·ЗіЈУРИӨөДПЦПуЈ¬ДЗҫНКЗјо»щЧйіЙПаН¬Ј¬ө«аСЯКәНаЧаӨөДЕЕБРЛіРтІ»Н¬Ј¬Л«ВЭРэөДОИ¶ЁРФҫЯУРПФЦшөДІоТмЎЈАэИз5ЎдGc 3ЎдЎЎ3ЎдG 5ЎдәН5ЎдGC 3ЎдЎЎ3ЎдGC 5ЎдөДОИ¶ЁРФПаІоәЬҙуЈ¬З°ХЯөДОИ¶ЁРФФ¶ҙуУЪәуЎЈЛьГЗөДЗвјьКэДҝКЗПаН¬өДЈ¬ЛьГЗөДІоұрФЪУЪПаБЪјо»щЦ®јдөД¶СјҜБҰІ»Н¬ЎЈјҙҙУаСЯКөҪаЧаӨөД·ҪПтөДјо»щ¶СјҜЧчУГПФЦшөШҙуУЪН¬СщЧйіЙөДаЧаӨөҪаСЯК·ҪПтөДјо»щ¶СјҜЧчУГЎЈ(ХвАпөД·ҪПтҫНКЗіЈ№жөДҙУ5Ўд¶ЛөҪ3Ўд¶ЛөД·ҪПт)ЎЈХвКЗТтОӘЗ°ХЯөДаСЯК»·әНаЧаӨ»·ЦШөьГж»эҙуУЪәуХЯөДаЧаӨ»·әНаСЯК»·өДЦШөьГж»эЈ¬ХвФЪBРНDNAЦРИ·КЗИзҙЛЎЈ
ЎЎЎЎёщҫЭGotoh 1981ДкөДСРҫҝЈ¬К®ЦЦПаБЪ¶юәЛЬХЛб¶ФөДTmЦөИзұн15?ЛщКҫЈ¬өҘО»ОӘЎжЈ¬ЛщУГАлЧУЗҝ¶ИОӘ19.5mmol/l NaЈ«ЎЈ
ұн15Јӯ5ЎЎПаБЪ¶юәЛЬХЛб¶ФTmЦө
| ЎЎ | 3Ўд | ||||
| A | T | G | C | ||
| 5Ўд | A | 54.50 | 57.02 | 58.42 | 97.73 |
| T | 36.73 | 54.50 | 54.71 | 86.44 | |
| G | 86.44 | 97.73 | 85.97 | 136.12 | |
| C | 54.71 | 58.42 | 72.55 | 85.97 | |
ЎЎЎЎУЙұн15-5ҝЙТФҝҙөҪЈ¬5ЎдTA 3Ўд 3ЎдAT5ЎдөДTmЦөЧоөНЎЈФЪХжәЛЙъОпЦРЈ¬іЈҝЙТФФЪӘІ19өҪӘІ27өДО»ЦГЙПҝҙөҪТ»ёцҪРЧцTATAҝтөДҪб№№(УЦіЖHognessҝт)Ј¬ХвКЗRNAҫЫәПГёөДҪбәПО»өгЎЈФЪХвАпRNAҫЫәПГёәНУР№Шө°°ЧЦКТтЧУРОіЙЧӘВјЖрКјёҙәПОпЎЈ
ЎЎЎЎУЦИзЈ¬ЙъГьУР»ъМеСЎФсUAAЧчОӘЧоУРР§өДЦХЦ№ГЬВлЧУҫшІ»КЗЕјИ»өДЈ¬ТтОӘ64ёцИэБӘМеГЬВлЧУЦРЈ¬ЛьУл·ҙГЬВлЧУ(јЩ¶ЁУРөД»°)РОіЙөД»ҘІ№ІъОп5ЎдUAA3Ўд3ЎдAUU5ЎдөДTmЦөКЗЧоөНөДТ»ёцЈ¬јҙК№ФЪЙъАнОВ¶ИПВТІКЗІ»ОИ¶ЁөДЎЈөұіхУРИЛ»ЁБЛәЬ¶а№Ө·тИҘС°ХТТ»ёцІ»РҜҙш°ұ»щЛбөДЧЁ№©лДБҙЦХЦ№УГөДtRNAЈ¬ЖдКөІўІ»ҙжФЪХвЦЦtRNAЎЈлДБҙөДКН·ЕКЗУЙКН·ЕТтЧУRFФЪЖрЧчУГЎЈФЪИэЦЦЦХЦ№ГЬВлЧУЦРЈ¬UAGәНUGAіЈ»бОӘН»ұдРНөДtRNAОЮТеТЦЦЖЈ¬¶шUAAФтәЬЙЩ·ўЙъОЮТеТЦЦЖТІҝЙДЬҫНКЗХвёцөААнЎЈХвТІҫНЛөГчБЛОӘКІГҙФЪлДБҙЦХЦ№ҙҰіЈіЈ»біцПЦЛ«ЦШЦХЦ№ГЬВлЧУЎЈ




