



��(��)ϸ��ɫ����ϵ
����1926��Keilin�״�ʹ�÷ֹ⾵�۲�������輡��ʱ����������������չ��ף���˰�ϸ���ڵ��������ʶ���Ϊϸ��ɫ�ء�ϸ��ɫ����һ�ຬ����߲��������ɫ���ף����ڵݵ����塣��������Ĥ����ϸ��ɫ��b��c1��c��aa3���Ρ�������֯����������ϸ��ɫ��P450��ϸ��ɫ��b��c1��cΪ��ɫϸ���أ�ϸ��ɫ��aa3Ϊ��ɫϸ���ء���ͬ��ϸ��ɫ�ؾ��в�ͬ�����չ��ף�������ø���ṹ��ͬ�������ĽṹҲ��һЩ���졣
����ϸ��ɫ��cΪһ���ܵ��ף�λ����������Ĥ����ࡣϸ��ɫ��C�Ƚ��������ᴿ����ṹ����������鶯���Cyt c��104���������л���ɣ����ӽ����ĽǶ����������о���Cyt c�ĸ���Ѫ����(����ԭ߲��)ͨ�����ۼ�(���Ѽ�)��ø��������(��ͼ6-2)���������ϸ��ɫ���и�����ø����ͨ���ǹ��ۼ���ϡ�
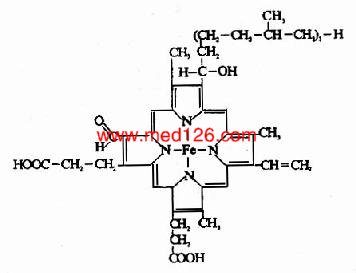
ͼ6��2��ϸ��ɫ��C�ĸ�����ø�������ӷ�ʽ
����ϸ��ɫ��a��a3���ֿ���ͳ��Ϊϸ��ɫ��aa3����ϸ��ɫ��P450��b��c1��c��ͬ��ϸ��ɫ��aa3�ĸ�������Ѫ���أ�����Ѫ����A(��ͼ6?)��ϸ��ɫ��aa3�ɽ�����ֱ�Ӵ��ݸ���������ֳ�Ϊϸ��ɫ������ø��
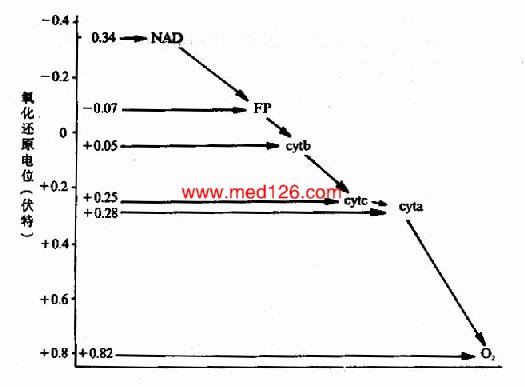
ͼ6-3��Ѫ����A�ṹʽ
������߲����������Fe2+����Fe2+����Fe3++e�Ļ��䣬������ݵ��ӵ����á���ԭ�ӿ��Ժ�ø����߲�����γ�6����λ����ϸ��ɫ��aa3��P��450�����е���ԭ��ֻ�γ�5����λ���������������γ�һ����λ����������ֱ�Ӵ��ݸ�����Ҳ����CO���軯�H2S������������γ�һ����λ����ϸ��ɫ��aa3���軯���Ͼ�����������������ĵ��Ӵ��ݣ������軯���ж���
���������������и��ִ����������˳��
����(һ)ȷ������˳��ķ���
����1.���ݸ�����ֵı�������ԭ��λ��ȷ������������ԭ��λ����ֵ��ʾ������ԭ�����Ĵ�С����������ԭ��λ��ֵԽ���仹ԭ��Խǿ�����ױ���������������ԭ��λ��ֵԽ����������Խǿ�����ױ���ԭ����˺������и�����ֵ�����˳��Ӧ���ɵ͵�λ������ߵ�λ����(ͼ��4)��
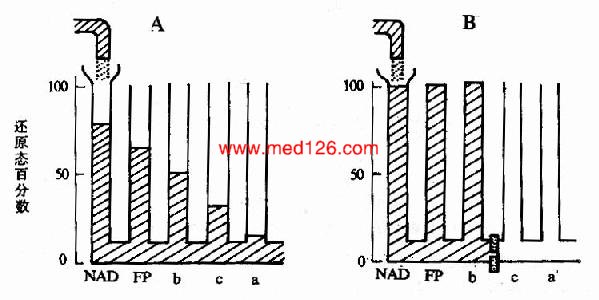
ͼ6-4�����ִ�����ı�������ԭ��λ
����2.����������������������Ӧ�ﵽƽ��ʱ���ִ�����Ļ�ԭ�̶���ȷ����Chance��Williamsʹ�÷ֹ��ȷ��ⶨ�����������������������������ѭ����Ӧ�ﵽƽ��ʱ���������и��ִ�����Ļ�ԭ�̶ȡ���Ӧ�ﵽƽ��ʱ�ӵ���һ�ൽ��һ��ĸ��ִ�����Ļ�ԭ�̶�Ӧ���ǵݼ��ģ������һ����ߣ���һ����ͣ����±���������ʾ����
��6-1��������̬ƽ��ʱ���Ӵ�����Ļ�ԭ�̶�
| ������ | NAD | FP | Cyt b | Cyt c | Cty aa3 |
| ��ԭ��% | 53 | 20 | 16 | 6 | 1 |
����FP�����ص���
�������������������ѧ�ϵ���ͨ�ܣ�ͼ6?A�У�����ˮ�����ڳ�ˮ�����������ﵽƽ��ʱ�����ˮ�������ˮ����ˮλ��ߣ����ˮ�������ˮ����ˮλ��ͣ��ӽ�ˮ�ܵ���ˮ��ˮλ���ͣ�����ˮ����Ϊ����������������ʵ���е������
����3.ʹ����������Ƽ�����������Ƽ�����Ϻ������е��ض����ڣ���ϲ�λ�ĵ���һ��ĸ��ִ�����ӦΪ��ԭ�ͣ���ϲ�λ����һ��ĸ��ִ�����ӦΪ�����ͣ��������������ͨ�ܵĵײ�һ������ϲ�λ��ǰ�ĸ�ˮ����ˮ�����ģ�����ϲ�λ�Ժ�ĸ�ˮ����ˮ������(��ͼ6-5��B)����ҽѧȫ.������.վ.�ṩ
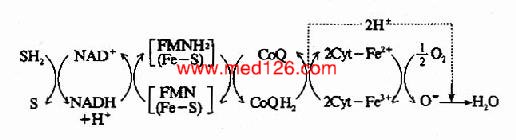
ͼ6-5�����������ȶ�ʱ���ִ�����Ļ�ԭ̫����
A.�������Ƽ���B.���뿹ùA���
�����������NADH������CoQ��ԭ��
�����������������������CoQ��ԭ
�����������Co QH2������Cyt c��ԭ
���������������Cyt c������O2��ԭ
��6-2��ʹ�ÿ�ù��Aǰ����ֵݵ�����Ļ�ԭ�Ͱٷ���
| FP | Cyt b | Cyt c+c1 | Cty aa3 | |
| ������ | 40 | 25 | 19 | 4 |
| �����ᣫ��ù��A | 100 | 100 | 0 | 0 |
�����ӱ��п��Կ�����FP��Cyt bλ�ڿ�ù��A��ϲ�λ֮ǰ��Cyt c��cl��aa3λ����ϲ�λ֮���ò�ͬ�����Ƽ�����ʵ�飬�Ϳ���ȷ���������и��ִ����������˳��
����4.������ʵ���У���������ֳɸ��ָ�����������Դ��ķ�Ӧ���ٽ������飬������������
������������(Green)��ʵ���ҳɹ��ؽ�����������ɾ��д����Ե����ָ������Լ�CoQ��Cytc.������������Ĺ��ܷ��֣�
�������Կ���CoQ�ڸ������������֮�䴫�ݻ�ԭ������Cyt c�ڸ���������֮�䴫�ݻ�ԭ�����������ֽ������ָ�����1��1��1��1�ı�����ϣ�����CoQ��Cyt c���飬�����ϻָ���������ԭ�еĴ�������
������������ʵ�鷽��������������ֵ�����˳���ѻ�����ȷ��������Щ��һ�µĿ�����������CoQ��ϸ��ɫ��C��һ�����о��û��ܲ����������Fe-S��CoQ�Ķ�λ������Ҳ�����顣




