



����(��)G����ż�������弰���ź�ת��
����G����ż������������������ʡ����༤�غ��������ӵ����壬��ζ�����Ӿ�������н�����Դ�������ص���������G����ż�������塣���������ڽṹ�Ͼ�Ϊ���嵰�ף�����ĩ��λ��ϸ������棬�Ȼ�ĩ���ڰ�Ĥ�ڲࡣ����������Ҫ������Ĥ�ߴ�(ͼ21-10)����������˽����������Ϊ�ߴο�Ĥ���塣��������������Ĥ����Ĥ����Ĥ�ڲ��γ��˼�����״�ṹ�����Ƿֱ���������(��ѧ�������ź�)�Ľ�Ϻ�ϸ���ڵ��źŴ��ݡ���������ֿ�����һ��GTP��ϵ���(���G����)����ã�����G�����Ǹ��źŴ���;���еĵ�һ���źŴ��ݷ��ӣ���Ҳ���������屻��ΪG����ż���������ԭ��
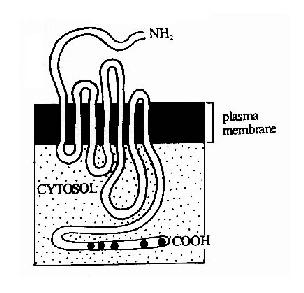
ͼ21��10���ߴο�Ĥ���壭G����ż��
���������Ĥ�ṹʾ��ͼ
����G����ż��������źŴ��ݹ��̰���(1)�����������ϣ�(2)����G���ף�(3)G�����������ϸ���е�ЧӦ���ӣ�(4)ЧӦ���Ӹı�ϸ������ʹ�ĺ�����ֲ���(5)ϸ������ʹ��������Ӧ�İз��ӣ��Ӷ��ı�ϸ���Ĵ�л���̼��������ȹ��ܡ����ڽ���һ������һ���̵���Ҫ���ڡ�
����1.G����ѭ����(G Protein Cycle)
����G����ż����������ź�ת��;���еĵ�һ���źŴ��ݷ�����G���ף������̳�ΪG����ѭ����
����G�����Ԧ����¦��ǻ����������ʽ������ϸ����Ĥ�ڲࡣ���ǻ��ѷ�����20���֣�������Ϊ36��52kd�����ǻ����ж���λ�㣬���а������������ϲ��������ڵIJ�λ����¦��ǻ����ϵIJ�λ��GDP��GTP��ϲ�λ�Լ�������ЧӦ��������õIJ�λ�ȵȡ����ǻ�������GTPø���ԡ����ǻ����GDPʱ������״̬������GTP���ʱ��Ϊ�л���״̬��GTP��ˮ����ʹ�䷵������״̬��
����G�����еĦºͦ��ǻ��������֣����������ǻ�����ࡣ��ϸ���ڣ��ºͦ��ǻ��γɽ��ܽ�ϵĶ����壬ֻ���ڵ��ױ��������·��ɽ��룬��˿�����Ϊ�����ǹ����ϵĵ��塣�¦��ǻ�����Ҫ����������ǻ��γɸ����岢��λ����Ĥ�ڲࡣ���������о��������¦��ǻ����������������ЧӦ���ӡ�

ͼ21��11��G����ѭ��ʾ��ͼ
����G����ѭ���ľ�����̿ɼ�ͼ21-11����������ѧ�źŴ̼�����ʱ������G����ʹ֮��������ı䡣���ǻ���GDP�������½�����ϵ�GDPΪGTP��ȡ�������ǻ������GTP����¦��ǻ��������룬��Ϊ�״̬�Ħ��ǻ�����˵Ħ��ǻ���ʱ�������������εĸ���ЧӦ���ӡ����ֻ״̬��һֱ������GTP�����ǻ��������е�GTPøˮ��ΪGDP��һ������GTP��ˮ�⣬���ǻ����ٴ���¦��ǻ��γɸ����壬�ص���ֹ״̬�����½����µĻ�ѧ�źš�
��������G�������ͬ�����G�����������ڲ�ͬ��ЧӦ���ӣ����ͬһЧӦ���ӽ��в�ͬ�ĵ��ڡ�




