



��������YEEI��src family��SH2���
����YMXM��growth factor receptor��P13Kp85��SH2���
����YVIP��PLC�õ�SH2���
����YXNX��Grb2��SH2���
����B.SH3�ṹ����50��100����������ɣ��鵼�źŷ����븻��������ĵ����ӵĽ�ϣ����������븬����л����ڽ�������л������ɵĻ���������ء�
��������RKLPPRPSK��P13K������Ϊ9.1��M
����PALPPLPRY��P13K������Ϊ17��M
����C.PH�ṹ����100��120����������ɣ��书����δ��ȫȷ����Ŀǰ��֪����������֬�����PIP2��PIP3��IP3�Ƚ�ϡ�ͬʱҲ���֣�һЩ�����ӣ���PKC��G���Ħ¦��ǵ�λҲ������PH�ṹ���ϡ�
����D.PTB�ṹ����Լ160����������ɣ���SH2һ����PTB�ṹ��Ҳ����ʶ��һЩ�������Ұ���Ļ������ϻ�����SH2�ṹ���������
������Ϊ���ؽ��Ԫ���������ڽṹ�������������ص㣺
������һ���źŷ��ӿ��Ժ����������ϵĵ��ؽ��Ԫ��(ͼ21-23)����˿���ͬʱ���������ϵ������źŷ������ϣ����磬�ڵ����Ұ��ἤøBtk�м���PH�ṹ��SH3�ṹ���SH2�ṹ���3�����ؽ��Ԫ����

ͼ21-23���ź�ת�������еĵ��ؽ��Ԫ����
����Y-Kinase��tyrosine kinaseDBD:DNA binding domain��PP:Pro��rich
����ABD:actin��binding domainGAP:��GTPase��activating domain
������ͬһ����ؽ��Ԫ���ɴ����ڶ��ֲ�ͬ���ź�ת�������У����磬PH�ṹ�������ijЩ����ø���ͷ�����G�����ڷ��Ӽ�ϸ���Ǽܵ��ȶ����ź�ת�������С���Щ���ؽ��Ԫ����һ���ṹ��Ȼ�Dz�ͬ�ģ���˶�����ϵ��źŷ��Ӿ���ѡ���ԣ����DZ�֤�źŷ�������þ��������ԵĻ�����
��������Щ�ṹ������Ϊ�Ǵ��ṹ��
����2.�������͵��ź�ת��;��
����(1)��Ƥ������������鵼���ź�ת��;��
������Ƥ���������������壭��Ƥ�������������Ϻ������һϵ��ϸ���ڱ仯������ʹϸ�������ֻ�����ֳ����Ƥ��������������һ�������Ұ��ᵰ��ø���������Ұ��ᵰ��ø��Ras��MAPK����;���DZ�Ƥ�������Ӵ̼��źŴ��ݵ�ϸ�����ڵ�����Ҫ;�����������³�Ա��ɣ���Ƥ�����������������SH2�ṹ��Ľ�ͷ����(��Grb2)�������ʺ������ͷ�����(��SOS)��Ras���ס�MAPKKK(��Raf1)��MAPKK��MAPK��ת¼���ӵ�(ͼ21��24)��
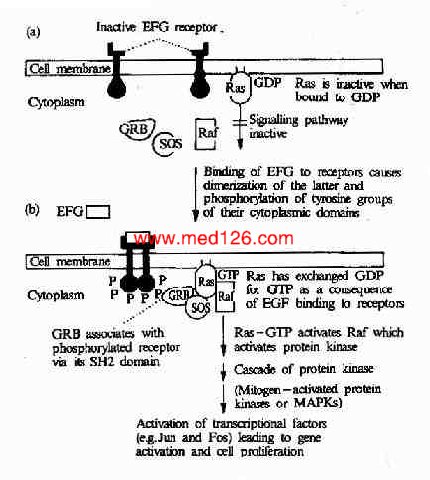
ͼ21-24��EGF����鵼���ź�ת������
������Ƥ���������������Ϻ���ʹ���巢�������廯���Ӷ��ı�������Ĺ���ʹ���еĵ����Ұ��ἤø������ǿ�������������Ұ���л��������ữ�����ữ��������γ����뺬SH2�ṹ��ĵ�����Grb2��ϵ�λ�㣬����Grb2������Ľ�ϡ�Grb2��������SH3�ṹ�ò�λ��һ�ֳ�ΪSOS�������ύ�����ӽ�ϣ�ʹ֮���Ըı䣬SOS���һ���Ras�������Ras������MAPK����ϵͳ������ERK�ļ�����ERKת�Ƶ�ϸ�����ڣ�����ijЩת¼���ӵĻ��Ըı�Ӷ��ı����ı���״̬��ϸ������ֳ��ֻ����̡�
����(2)�ã�����������鵼���ź�ת��
�����ã����������ɻTϸ�������ģ������дٽ���ԭ��ʺ�����������ʶ������ã����ɴٽ�Bϸ�����ڿ��塣�ã����������������Ժ�Ҳ���Ե�����������廯�������廯��������Լ���JAK��STATϵͳ�����߽������ش̼��źŴ�����ڡ�JAK(Janus Kinase)Ϊһ�ִ����ڰ����еĵ����Ұ��ἤø��������ʹ�������������ữ��STAT(Signal Transducerand Activator of Transcription)����ͨ����SH2�ṹ��ʶ�����ữ�����岢��֮��ϡ�Ȼ��STAT��������Ұ�������ữ���Ұ������ữ��STAT��������γ��л��Ե�ת¼���ӣ�Ӱ�����ı���(ͼ21��25)��

ͼ21-25���ã�����������鵼���ź�ת������
����JAK��Janus kinase
����STAT: Signal Transducer and Activator of Transcription
����GAS: �ê�interferon��activated sequence element
�������������ź�ת��;�������Ƕ����ź�ת��;���Ĵ���������90���������ѧ������ϸ���ź�ת���ķ��ӻ����о������Ѿ�ȡ����һЩ�ɾͣ����������ϸ���д��ڵ�ȫ����������ϵͳ��ʮ��ңԶ���д���ѧ���Dz���Ŭ�������¸�����ʵ��������ʶ���ҵ�Ը����




