



�����ġ�����̽��ı�Ǻͼ�⣨���ÿʮ���¡�)
���������ӽ��Ǻ�����������Թ����һ�ֽ�Ϸ�ʽ���Ǻ������Ҫ�������ԡ����÷����ӽ���һ���������ض��������н��м�⣬���뽫�ӽ����е�һ����ij�ֿ��Լ��ķ��ӽ��б�ǣ��������ͳ�Ϊ����̽�롣��ˣ�����̽����Ʊ��Ƿ����ӽ������Ĺؼ���������õ�Ҳ��Ŀǰ��õĺ���̽���Ƿ����Ƿ�����ͬλ�ر�ǡ����õķ�����ͬλ����32P��35Sǰ�������ߣ��ź�ǿ����á�������ͬλ�ر��̽����Ȼ���жȸߣ���ȴ���ڷ���Σ���Ͱ�˥�����ƣ�32P��˥��Ϊ14.3�죬35S��˥��Ϊ87.1�죬125I�İ�˥��Ϊ60��)��3H�İ�˥�ڳ���12.3�꣬�������ͷŦ·���������̫�ͣ�0.018Mev)��ֻ��������֯ԭλ�ӽ�������ͬλ�ر�ǵ�̽����ʹ�ù����д���������ȱ�㣬��Щ������������Ѱ�ҷǺ����Ա���﷽��ȡ���˺ܴ��չ�����������ж�ҹ�˾����Ƴ����ַǷ�����̽�����Լ��У��ڹ���Ҳ�Ѿ߱������������������������������Ӧ�Լ����ۡ�Ŀǰ���Ƿ����Ա�������������ࣺ������Hg��ӫ��������FITC���뿹ԭ���ظ����������أ�ø����������������ø��HRP)����������ø���������ø��AKP)�ȡ���ͬ�ı��������̽��ķ�������ⷽ��Ҳ���졣����������Ͻϳ��õģ���ʵ�ü�ֵ��չǰ���ļ��ֺ����Ƿ���������ʾ������������������¡�
������һ)����̽��ij���ø�ٱ�Ǽ���
����1��ȱ��ƽ�ơ��ü�����Kelly����1970�괴������ԭ����������DNAø��˫��DNA̽����ӵ�һ����������һЩȱ�ڣ�nick ),ȱ�ڴ����γ�3�����ǻ�ĩ�ˣ���ʱ���ڴ˾�DNA�ۺ�øI�Ĵ��½�������л�����3�����ǻ��ϣ�ͬʱ�����ݴ˾�øDNA�ۺ�øI��5����3����������ø���ԣ���ø��ȱ��5��������������г�����������ȱ��ƽ�ƣ�nick tr��anslation)���������ԭ��������ø�ǿ�ȵķ����Ժ����ᣨͨ��Ϊ��-32PdATP)�û���ǰ���ڵĺ����ᣬ����Ʊ��Ȼ��Ըߴ�108cpm��ÿ���Ӽ���)/��g��32P���DNA����ȱ��ƽ�Ʒ���ǵ�DNA̽�������������ӽ�Ҫ��
����2��DNA����ĩ�˱�ǡ��˾�DNA�ۺ�øi ���ݲݸ˾�����ø�и�ɵõ����������������з�����Ϊ76kd�Ĵ�Ƭ�γ�ΪKlenowƬ�Ρ���ø���������ۺ�øI��5����3���ۺ�ø���Ժ�3����5����������ø���ԣ���ȱ��5����3����������ø���ԡ�����KlenowƬ�ο����������ø����DNA��������3������ĩ�ˡ���ˣ������ַ������Ա��˫��DNA�İ���3��ĩ�ˡ���KlenowƬ�α��ĩ��һ��ֻ��һ��[��-32P]dNTP�����뷴Ӧ��[��-32P]dNTPȡ����DNAĩ�������5��ĩ�����У����磬��Ecor I�и�DNA��������ĩ����[��-32P]dNTP��ǡ���Ƿ�Ӧ����һ������ø����DNA���������У�����ȥ������ø��ʹ��ʧ�Ҳ�����������Һ������3�������DNAĩ�˲��ܱ�KlenowƬ����Ч�ڱ�ǣ������������ӿ���T4DNA�ۺ�ø��
����ѡ�����ֱ�Ƿ�����Ϊ�˲���������������Ӿʱ����С�������DNAƬ�Ρ���Ϊ��ǵ�DNAƬ������Ħ��Ũ�ȳɱ�����������Ƭ�δ�С�ɱ�����������ø�������У�С�ĺʹ��Ƭ�ζ�����ͬ�̶ȱ���ǡ���ˣ���ʹ�÷�������Ӱ��ȷ�����б��廯�Ҷ�Ⱦɫ���۲쵽�Ĵ�СDNA��������������Southern��ӡ�ӽ�ʱ��������־��ı�ǡ�ͨ��ѡ����Ӧ��ǵ�dNTP���÷�������ֻ���DNA���ӵ�һ�ˡ����磬��DNAƬ�ε�����ĩ�˷ֱ���Bam H I��Hind III ճĤ�ˣ��ڷ�Ӧ��ֻ����[��-32P]dNTP��[��-32P]dGTP��ʹ��ѡ���Ա����ĩ��֮һ��
����3����T4������ἤø���DNa 5��ĩ�ˡ��Ѻ�����̽���̵�RNA��DNA̽���ѡ�ô˷���ǣ��Ѻ�����̽��һ��������ֱ�ǡ�T4������ἤø��polynucleotide kinase, PNK)����T4�ɾ����Ⱦ�Ĵ˾�����ȡ�ģ���ø�ܴ�ATP�Ħ�-����ת����DNA��RNA��5����OHĩ�ˡ��ڹ���ADP����ʱ��Ҳ�ɴٽ����ύ����Ӧ��ʹPNK��DNAĩ��5������ת�Ƶ�ADP������ATP��Ȼ���[��-32P]dNTP�ϵı������ת����DNA��5��ĩ�ˣ��Ӷ�ʹDNA�������ữ������˵õ���ǡ���Ȼ��PNK���DNAĩ����Ҫ[��-32P]dNTP������ǰ��ø�ٱ�Ƿ�����ͬ��ͨ��������5�����ữ��DNA̽�룬Ҫ���ü�������øȥ��������ţ�Ȼ��������PNK����5��ĩ�˱�ǣ��������Ч�ʽϸߡ�
����4������������졡�����Ե���DNA��RNAģ��ϳɸ߱Ȼ���32P���̽����ѡ�õķ�����ԭ����ʹ��6��8nt�ĹѺ�����Ƭ������Ե�DNA��RNAģ���˻���DNA�ۺ�øI��ת¼ø�������£���ÿһ���˻�ģ���ϵĹѺ�����Ƭ��Ϊ��������DNA���ĺϳɣ��ڷ�Ӧʱ��[��-32P]dNTP����ϳ��������õ���ǡ����Դ������ºϳ�����̽��Ƭ��)��ģ����룬���õ��������ִ�С��̽��DNA����Ϊ���ùѺ�����Ƭ�κ̣ܶ��ڵ��������¿���ģ��DNA��������˻�Ӧ����˱���Ϊ������random primer)����������������Сţ����DNA���㾫DNA�Ʊ���
��������������ǵ�DNA̽���cDNA̽��Ȼ�����������ȱ��ƽ�Ʒ����ҽ����Ϊ�ȶ������ַ����������������DNA̽�룬��Ϊ��������������DNA������������е��˻���Ҫ����ԭ�����С���ˣ����ڿ�¡��DAN̽�룬���Ƚ�����̽��DNA���������պ��ٱ�ǣ���ȱ��ƽ�Ʒ���ֱ������ȫ�����ı�ǡ�
����5���ۺ�ø����Ӧ������ڶ�ʮ����)���ۺ�ø����Ӧ��polymerase chain reaction, PCR)��һ�ַ�������ѧ�¼�����������Cetus��˾�����Ŵ�ѧ����Kary.B. Mullis��1985�괴�����ü��������������෴���ӽ��������ڰ�DNA����ĹѺ��������ᆳø�ٺϳ������DNAƬ�Σ�����ģ����ԣ������˻������������������ķ���ѭ�����������������а�DNA�õ�ǧ�����ϵ���������� ��PCR�����ѳ�Ϊһ�Ϊ�м�ֵ�ļ�������Ѹ���ƹ�Ӧ�á�
����PCR������������Ҫ��;������֮һ���ǿ�������Ǹ߱Ȼ���DNA̽�롣PCR�������кܸߵ������ԣ�����1��2h֮�������ϳ�̽��DNAƬ�Σ�����ڵ����м���[��-32P]dNTP��������ǵ�dNTP����̽��DNA�ϳɹ����пɵõ��ܺõı�ǣ������IJ����ʿɸߴ�70%��80%����ˣ�PCR��Ǽ����ر������ڴ��ģ���ͷǷ����Ա�ǡ��÷���ȱ����Ҫ�ϳ�һ��������PCR���ʹ�ô�̽��DNA���Ʊ���СƬ��������Ҳ��ȡ�ýϺõı��Ч����
��������)����̽��ķǷ����Ա�Ǽ���
����1�����������ر�Ǻ��ᡡĿǰʹ�õĹ���������Լ������֣��������أ�������)������֬�������ء����Ƕ�����һ���������š�һ�����ӱۺ�һ�������ػ�����ɡ��������صĹ���������-N3�����ڹ������¿�������еļ����ϡ�����֬���������еĹ������Ų���֬���ڹ��գ�320��400��m)�£����뵥����˫�����ᷢ����Ӧ����Ӧ��Ҫ��T�ϣ�C��Ҳ��һ���̶ȵķ�Ӧ�����������ص����ӱۺ�6��12��̼ԭ�ӣ����Լ���̽���ӽ�ʱ�Ŀռ�λ�衣���������ر�Ǻ��ᣬ������������Ҳ�ɴﵽpgˮƽ����������Դ����ļ�⡣���������һ���µĹ�������DNA�������Լ�����������-���Ҷ���-�����أ�BPA)��BPA��DNA��ϲ�����һ���������㶹��������ڳ���UV��������DNA������ۼ���ϡ�BPA��Ӧ����DNA��ϱȹ��������ظ����죬�ڿɼ�������������ᷴӦ��������Կ�ʹBPAֻ��Ǵ���ϸ���ѽ����еĺ��ᣬ������ǵ��ס����Ǻ�����ϸ������ӡ�
����2��ø�������ر�Ǻ��ᡡ�������ػ����������������ᣨBio-11-dUTP,Bio �C7 �C dATP��Bio-11-dCTP)�ȴ�����Ӧ32P��ǵ��������������ᣬ��DNA�ۺ�ø���ò���DNA��Bio-dUTP����dTTP,Bio-dATP����dATP��Bio- dCTP����dCTP��Bio �C11-dUTP��11��ָ�����ػ���������������֮�����ӱ۵�̼�����ȡ����õ�ø�������ر��DNA�ķ�����ȱ��ƽ�Ʒ�������������취��
����3���Ѻ������������ĩ�˱�ǡ���5��-����Ļ�ѧ��Ƿ���3��-OH��ø�ٱ�Ƿ���ǰ�߽��Ѻ������5��-�������һ���Ҷ�����Ȼ�����������ǰ������أ��������ػ��������������������ϡ���������ĩ��ת�ƽ�Bio-11�CdUTP������3��-OH�ˣ���ȥһ��������)��ҽѧȫ����www.med126.com
����4��ø��DNA������Լ�������������ø��HRP)���������ø��AP)��ͨ���Ա�����PBQ)�����ϩ�ǰ���PEI)���Ӷ��ɣ�HRP-PBQ-PEI+)�����Լ������ȩ������������Ե�DNA��ϣ�ʹHRP��DNA������һ�����HRP��ǵ�DNA̽�롣
����5��ø��Ѻ����ᡡ����������5��ĩ�˱��HRP�����ڲ����AP����ǰ������HRP�в���һ��-HS��Ӧ���ţ��ڹѺ�����ϳ����˼���5���ˣ���һ��C6��-HS��������HRP��Ӧ����5��-HRP�Ѻ����ᡣ��������ȫ�ɹѺ���������н�һ��5�������ӱۼ�CF3���ŵ�����3���������ǰ��ϳ��ڹѺ��������У��ϳɺ�˻�ĹѺ�������AP��Ӧ���õ�AP��ǵĹѺ����ᡣ
����6��DNA�뿹ԭ��ǡ���ԭ����Bio-11-dTUP��ͬ��ֻ����ë�ػ�߰�����������γ�Dig �C11- dUTP��ø�ٲ���DNA���ӡ��ÿ�ë�ػ�߰����������DNA�ϵİ뿹ԭ����Digoxigenin���ظ���)
����(��)�Ƿ�����̽�����ʾ��ϵ
����1��AP��ɫ��ϵ

����BCI �COH+NBT����ɫ��
����ASO����λ��������ĹѺ����ᣬBCIP��5��-4��-3�������ᣬNBT���ĵ�������Pi�����ᡣ
����2��HRP��ɫ��ϵ
����HRP + H2O2[HRP��H2O2]
����ODA �CNH2 +[HRP ��H2O2]����ODA-N = ODA/��ɫ��+HRP +H2O
����ODA����-����������
����3��ABC��ɫ��ϵ
����DNA �CB + SA �C AP��DNa �C B �C SA��DNa �C B +SA - BAP��DNa �CB �CSA �CBAP
��������������Ӧ����AP������DNA���Ժ��ٽ���APø��ɫ������SAΪStreptavidin����ø����)��BAPΪ�����ػ�������ø��BΪ�����أ�biotin)��ABCΪAvidin �C Biotin - Enzyme com��plex, ������- ������-ø�����
�������Ϸ�ӦAP�����HRP���档
��18-3������̽��ı�Ƿ���
| ��������� | ��Ƿ��� | ��Ƿ��� | �ӽ���ļ�ⷨ |
| �����Է��� | [��-32P]dNTP | NT��PCR��RPI | ��������Ӱ����� |
| �� | [��-32P]dNTP | TL | ��������Ӱ����� |
| �� | 125I | �⻯ | ��������Ӱ����� |
| �� | 35S | NT | ��������Ӱ����� |
| �� | 3H | NT | ��������Ӱ����� |
| �Ƿ����Է��� | �� | �� | �� |
| ������ | Bio-11-dUTP | NT��TL��PCR | ø�����ػ�ø�꿹�����ؿ�����ɫ |
| �� | ���������� | 600W �ɼ����� | ͬ�� |
| �� | �����ػ�����֬�� | 365nm�������� | ͬ�� |
| �� | ��������-e-�������� | ��ѧ�ϳɷ� | ͬ�� |
| �� | N-�ǻ��������ǰ�֬ | �� | ø�����ػ�ø�꿹�����ؿ�����ɫ |
| �� | �������� | ��ѧ�ϳɷ� | ø�����ػ�ø�꿹�����ؿ�����ɫ |
| ø | ��������ø | ֱ�ӷ���ϳɷ� | ֱ�ӵ�����ɫ����ø�꿹��+ ������ɫ |
| �� | ����������ø | ֱ�ӷ���ϳɷ� | ͬ�� |
| ӫ���� | ������FITC | �ϳɷ� | ֱ��ӫ�������۲��ø�꿹��+ ������ɫ |
| ��ѧ�������� | �� | �� | ��ѧ�������������Ӱ |
| ��˫������ | ø���ӫ���ǵ��� | ��ѧ�� | ֱ�ӵ�����ɫ |
| ϡ�н��� | Eu3�� | ��ѧ�� | ʱ��ֱ�ӫ�� |
| �ؽ��� | Ag | ��ѧ�� | �� |
| �� | Hg | ��ѧ�� | �� |
| �뿹ԭ | �������� | ��ѧ�� | ø�굥�� + ������ɫ |
| �� | �ظ��� | RPL | ø�꿹�� + ������ɫ |
����* ��NT��ȱ��ƽ�ƣ� PCR���ۺ�ø����Ӧ��RPL�� ��������ǣ� TL��ĩ�˱��
����4���Ƿ��䷢������Ӱ��������AP��HRP����ɫ������ݹ⻯ѧԭ������һ��ø���������ӵĻ������������Ӱ��������X��Ƭ��ʾ��
������1)HRP��������Ӱ��������-��������HRP��H2O2������������Ϊ�����������ᣬͬʱ�ų�N2�����⣨����428nm)������ʱ����ijЩ�ӵ�������ʱ����ǿ������ǧ������Ӧ����ͼ��
������2)AP�ķ�������Ӱ��AP�ķ�������ǽ����������������Σ�AMPPD)������������������AP��������ˮ����һ�����ᣬ�����ɷ����ڹ������ṩ��Դ�ֽ�������ͪ�ͼ���̬�ļ���-�������������ӣ����������ӻָ�����̬ʱ�������ӡ����ò����ڰ�Ƭ��621��)ֱ�ӱ�����Ӱ����Ӱ�ź�ǿ�ȱ�BCIP/NBP��ɫ��ǿ�������������Ǻ�ǰ������ʾ��ϵ��
�����䷢�ⷴӦ��ԭ�����£�
����HRP�ķ���ԭ����
����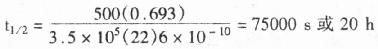
����AP�ķ���ԭ����
����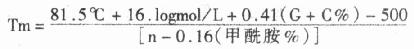
��




