



C5是形成膜攻击复合体(MAC)的第1个补体分子。C5由以二硫键相连接的α、β链组成,分子量190 kDa,其中α链为115kDa,β链为75kDa(图5-9)。C5与C3和C4的结构相类似,但没有链内硫酯键。靠近N端的第74-75位精氨酸一亮氨酸键为C5转化酶作用的部位。在C5转化酶的作用下,C5α链N末端裂解出一个分子量为11kDa的小片段C5a进入液相中,其余部分为110kDa的大片段C5b,仍结合在细胞膜表面。亲生的C5b在极短时间内能保持与C6结合的构象,可与C6非共价结合形成一牢固的C5b6复合物,并通过与C3b的可逆性结合而固定的细胞膜上。但C5b生成后其潜在的生物学活性存在时间非常短促,若无C6结合则迅速衰变为C5bi。
C5b只形成MAC参与细胞溶解效应,而C5a却具有广泛的生物学活性。概括起来有以下几方面:(1)过敏毒素作用:C5a是具有过敏毒素作用的补体裂解片段中作用最强的介质,较C3a强20倍,较C4a强2500倍。此外,C5a还可不依赖于肥大细胞释放组胺,即通过直接作用于血管内皮细胞而增加血管的通透性。(2)趋化作用:高浓度的C5a是中性粒细胞、嗜酸性粒细胞和单核细胞的趋化剂,可刺激这些细胞沿着浓度定向移动。值得注意的是,被血清羧肽酶N切C5a C端精氨酸残基而形成的去精C5a 虽丧失了使肥大细胞分泌组胺的能力,但仍具有较强的趋化活性,是补体活化后产生趋化作用的主要因素。(3)促代谢作用:高浓度的C5a可刺激中性粒细胞和单核细胞的氧化代谢,提高其cGMP的水平,有利于促进溶酶体与细胞膜的融合,释放溶酶体酶。此外,C5a还可刺激中性粒细胞粘附及增强其产生超氧化物。(4)免疫调节作用:近年体外研究表明,C5a对免疫应答有明显增强作用,如可诱导单核细胞分泌IL-1、IL-6、IL-8及TNF-α等细胞因子,促进抗原及同种异体抗原诱导的T细胞增殖及B细胞产生抗体等。C5a的上述生物学活性的利于增强机体的防御机能,但由其导致的炎症反应也可造成对机体的损伤。编码入C5的基因定位于第9号染色体长臂32-34区。医学全在.线www.lindalemus.com
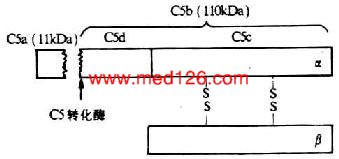
图5-9 C5分子的结构(模式图)




