



(二)CD3(T3、Leu4)
CD3分子分布于成熟T淋巴细胞表面,至少由γ、δ、ε、ζ、η5种多肽链组成,与T细胞抗原受体非共价连接(图1-3)。CD3单克隆抗体可诱导CD3多肽和TCR共帽形成(co-capping),并诱导T淋巴细胞活化。TCR识别外来抗原与自身MHC分子形成的复合物,CD3对于信号的传递具有重要作用。
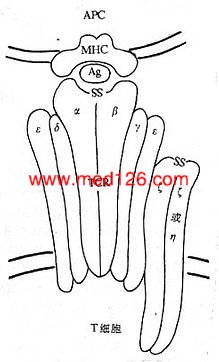
图1-3 TCR/CD3结构模式图
T细胞在胸腺发育过程中,CD3γ、δ和ε基因的表达要早于TCRα、β链基因的表达。CD3γ、δ和ε基因产物通过翻译后的修饰形成核心结构,在内织网处,此核心结构与TCRαβ异源双体形成复合物后转移到高尔基氏体,进行N连接的糖基化。ζ-ζ同源双体与TCRαβ/CD3γδεε结合后组成一个完整的复合物TCRαβ/CD3γδεεζζ(少娄分子为TCRαβ/CD3γδεεζη)。最近发现一个分子量为28kDa的ω连或T细胞受体相关蛋白(t cell receptor associated protein,TRAP),可能具有控制TCR/CD3复合物在内织网中装配和转移的功能,但确切的机理尚不明了。ω链不表达于细胞膜表面。
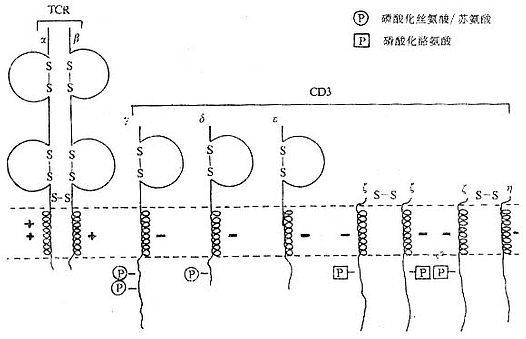
图1-4 TCR/CD3复合物模式图
1.CD3γ、δ和ε链 CD3γ、δ和ε链基因有高度的同源性,在人类位于第11号染色体,小鼠9号染色体,这三种链的基因可能从一个祖先基因通过基因复制而来。CD3γ、δ和ε链在细胞膜外都有一个Ig样结构域(C2),都属于免疫球蛋白超家族,但不存在多态性或可变性,因此不直接参与特异性识别抗原。γ、δ和ε链的穿膜部分含有带负电谷氨酸和/或天冬氨酸残基,这与TCRα、δ链穿膜区中带正电赖氨酸、精氨酸以及β、γ链穿膜区中的赖氨酸相互作用有重要作用。γ、δ和ε链胞浆部分含44到81氨基酸残基,提供了把信息传导到细胞内的条件。γ链分子量为25~28kKa,有2个糖基化点,氨基端89个氨基酸残基为亲水性,组成胞膜外区,穿膜区含27个氨基酸残基,胞浆内区44氨基酸残基,胞浆内113位丝氨酸残基可能是磷酸化位点。δ链分子量为20kDa,含有2个糖基化点,胞浆内126位丝氨酸可能是磷酸化位点。CD3δ链抗体能非特异性地活化T细胞,促进T细胞有丝分裂。ε链分子量为20kDa包括氨基端104亲水氨基酸的胞膜外区,穿膜区26个氨基酸残基,胞浆内区81个氨基酸残基。目前所制备的单克隆抗体中大部分是针对CD3ε链。
2.CD3ζ和η ζ(zeta)和η(eta)链结构相似,而与CD3γ、δ和ε链无同源性。ζ和η链分子量分别为16kDa和21kDa,它们的胞膜外以及穿膜区和结构相似,但有胞浆区不同。胞膜外区很短,只有9个氨基酸残基,含有半胱氨酸,ζζ之间或ζη之间形成二硫键。ζ和η链穿膜部分各有一个带负电的天冬氨酸。ζ和η链胞浆内区分别有113个和155个氨基酸残基,具有多个酪氨酸磷酸化的位点。最近研究证实,CD3ζ链可能与NK细胞上Fcγ受体相连。此外,ζ链与FcεRⅠγ亚单位有很高的同源性。
CD3γ、δ和ε链是单链,而ζ则以ζ-ζ同源双体存在于80~90%T细胞中,有10~20%T细胞则以ζ-η异源双体存在。因此最常见的TCR/CD3复合物的组成形式是TCRαβ/CD3γδεεζζ。
在体外,抗CD3McAb可促进T细胞表达IL-2R,产生IL-2、TNF-α、TNE-β、IFN-γ和IL-4等多种细胞因子,诱导非MHC限制的细胞毒作用,增强T细胞、LAK和NK细胞的杀伤肿瘤作用。
1986年美国FDA已批准应用小鼠抗CD3 McAb治疗急性肾移植排斥反应。CD3McAb治疗心、肝移植排斥反应已完成Ⅲ期临床试验,正申请投放市场。




