



三、第二信使
受体在识别相应配体并与之结合后需要细胞内第二信使(second messenger) 将获得信息增强、分化、整合并传递给效应机制才能发挥其特定的生理功能或药理效应。最早发现的第二信使是环磷腺苷(cAMP),现在知道还有许多其他物质参与细胞内信息转导。这是一个非常复杂的系统,简示如下(图2-14),很多问题尚有待进一步阐明。
1.G-蛋白 G蛋白是一类存在于细胞膜内侧的调节蛋白,都是由三个不同亚单位α、β、γ组成的三聚体。静息状态时与GDP结合。相应受体激活后GDP-α、β、γ复合物在Mg2+参与下,结合的GDP与胞浆中GTP交换,GTP-α与β、γ分离并与相应的效应机制结合,同时配体与受体分离。α亚单位内在的GTP酶活性促使GTP水解为GDP,激活效应机制,从而恢复原来静息状态(图2-15)。GS激活腺苷酸环化酶(AC),使cAMP增加。Gi抑制AC,使cAMP减少,G-蛋白还激活磷脂酶C(PLC),调节Ca2+、K+等离子通道。对鸟苷酸环化酶也有激活作用,作用非常广泛,介导多种效应。近来发现G-蛋白还介导激活磷脂酶A2(PLA2)而产生花生四烯酸(AA),后者是各种前列腺素及白三烯的前体。
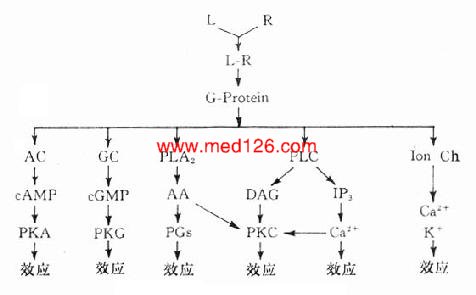
图2-14 第二信使系统示意图
2. 环磷腺苷(cAMP) cAMP是ATP经AC作用的产物。β受体、D1受体、H2受体等激动药通过GS作用使AC活化,ATP水解而使细胞内cAMP增加。α受体、D2受体、MACh受体、阿片受体等激动药通过Gi作用抑制AC,细胞内cAMP减少。cAMP受磷酸二酯酶(phosphodiesterase,PDE)水解为5’AMP后灭活。茶碱抑制PDE而使胞内cAMP增多。cAMP能激活蛋白激酶a (PKA)而使胞内许多蛋白酶磷酸化(ATP提供磷酸基)而活化,例如磷酸化酶、脂酶、糖原合成酶等活化而产生能量。钙离子通道磷酸化后激活,钙离子内流而使神经、心肌、平滑肌等兴奋。

图2-15 G-蛋白作用示意图
3.环磷鸟苷(cGMP) cGMP是GTP经鸟苷酸环化酶(GC)作用的产物,也受PDE灭活。cGMP作用与cAMP相反,使心脏抑制、血管舒张、肠腺分泌等。CGMP可以独立作用而不受cGMP制约。cGMP可激活蛋白酶G而引起各种效应。
4.肌醇磷脂(phosphatidylinositol) 细胞膜肌醇磷脂的水解是另一类重要的受体信息转导系统。α、H1、5-HT2、M1、M3等受体激动药与其受体结合后通过G-蛋白介导激活磷脂酶C(PLC)PLC使4,5-二磷酸肌醇磷脂(PIP2)水解为二酰甘油(DAG)及1,4,5-三磷酸肌醇(IP3)。DAG在细胞膜上激活蛋白激酶C(PKC),使许多靶蛋白磷酸化而产生效应,如腺体分泌,血小板聚集,中性粒细胞活化及细胞生长、代谢、分化等效应。IP3能促进细胞内钙池释放Ca2+,也有重要的生理意义。
5.钙离子 细胞内Ca2+浓度在1μmol/l以下,不到血浆Ca2+的0.1%,对细胞功能有着重要的调节作用,如肌肉收缩、腺体分泌、白细胞及血小板活化等。细胞内Ca2+可从细胞外经细胞膜上的钙离子通道流入,也可从细胞内肌浆网等钙池释放,两种途径互相促进。前者受膜电位、受体、G-蛋白,蛋白激酶A(PKA)等调控,后者受IP3作用而释放。细胞内Ca2+激活蛋白激酶C(PKC),与DAG有协同作用,共同促进其他信息传递蛋白及效应蛋白活化。很多药物通过对细胞内Ca2+影响而发挥其药理效应,故对细胞内Ca2+调控及其作用机制近年来受到极大的重视。
四、受体的调节
受体虽是遗传获得的固有蛋白,但并不是固定不变的,而经常代谢转换处于动态平衡状态,其数量,亲和力及效应力经常受到各种生理及药理因素的影响。连续用药后药效递减是常见的现象,一般称为耐受性(tolerance)、不应性(refractoriness)、快速耐受性(tachyphylaxis) 等。由于受体原因而产生的耐受性称为受体脱敏(receptor desensitization)。N2-ACh受体在受激动药连续作用后若干秒内发生脱敏现象,这是由于受体蛋白构象改变,钠离子通道不再开放所致。β-Adr受体脱敏时不能激活AC是因为受体与G-蛋白亲和力降低,或由于cAMP上升后引起PDE负反馈增加所致。具有酪氨酸激酶活性的受体可被细胞内吞(endocytosis)而数目减少,这一现象称为受体数目的向下调节(down regulation)。受体与不可逆拮抗药结合后其后果等于失去一部分受体,如银环蛇咬伤中毒时,N2-ACh受对激动药脱敏。与此相反,在连续应用拮抗药后受体会向上调节(up regulation),反应敏化。例如长期应用β-Adr受体拮抗药后,由于受体向上调节,突然停药时会出现反跳反应。




